Samenvatting
De influenza-epidemie van het seizoen 2018/2019 begon in week 50 van 2018 en duurde 14 weken. De epidemie piekte in week 6 van 2019 met een incidentie van 10,8 personen met influenza-achtige ziektebeelden per 10.000 inwoners. Influenza A(H1N1)pdm09- en A(H3N2)-virussen werden ongeveer even vaak gedetecteerd. Opvallend was de zeer sporadische detectie (minder dan 1 procent) van influenzavirus type B dit jaar. Ongeveer 81 procent van de circulerende A(H3N2)-virussen behoorde tot clade 3C.2a1b, waarvoor de antigene overeenkomst met de vaccinstam redelijk tot goed was. Er werd echter, net als elders, een groeiend aandeel (17 procent) clade 3C.3a A(H3N2)-virussen waargenomen, wat een probleem opleverde voor de vaccinkeuze voor volgend jaar. De A(H1N1)pdm09-virussen behoorden genetisch tot clade 6b.1a en vertoonden goede antigene gelijkenis met de vaccinstam. De vaccineffectiviteit tegen laboratoriumbevestigde influenzavirusinfecties was circa 57 procent. Onder de 756 virussen die zijn getest op gevoeligheid voor neuraminidaseremmers waren drie A(H1N1)pdm09-virussen met een sterk verlaagde gevoeligheid voor oseltamivir, in een geval in een onbehandelde patiënt. Alle circulerende influenza A-virussen die werden getest droegen een S31N aminozuursubstitutie in M2 die resistentie tegen ionkanaalblokkers bepaalt. Voor het seizoen 2019/2020 heeft de WHO voor het noordelijk halfrond de volgende vaccinsamenstelling aanbevolen:
- Voor A(H1N1)pdm09 een A/Brisbane/02/2018-achtig virus;
- Voor A(H3N2) een A/Kansas/14/2017-achtig virus; clade 3C.3a
- Voor de B/Victoria-lijn een B/Colorado/6/2017-achtig virus;
- Voor de B/Yamagata-lijn een B/Phuket/3073/2013-achtig virus.
Abstract
The influenza epidemic of 2018/2019 started in week 50 of 2018 and lasted 14 weeks. The epidemic reached peak incidence in week 6 of 2019, with 10,8 persons presenting influenza-like illness (ILI) per 10.000 inhabitants, marking a relatively mild season. Influenza A(H1N1)pdm09 and A(H3N2 viruses were detected at approximately equivalent frequencies. The sporadic detection (less than 1 per cent) of influenza B viruses was remarkable. Approximately 81 per cent of the circulating A(H3N2) viruses belonged to clade 3C.2a1b, for which the antigenic similarity to the vaccine strain was fair to good. Viruses of clade 3C.3a were detected with increasing frequency nationally (17 per cent) and elsewhere, causing a problem for vaccine strain selection. The A(H1N1)pdm09 viruses belonged to genetic clade 6b.1a and displayed good antigenic similarity to the vaccine strain. The vaccine effectiveness against laboratory-confirmed influenza virus infection was 57 per cent. Among 756 viruses tested for sensitivity to neuraminidase inhibitors, three had strongly reduced sensitivity. All tested influenza A viruses had the S31N amino acid substitution in M2 responsible for resistance to ionchannel inhibitors.
Griep
Griep wordt bij mensen veroorzaakt door influenzavirussen van het type A, B of C.[1] Sinds 2009 worden griepepidemieën wereldwijd afwisselend gedomineerd door twee influenzavirussen van type A en twee influenzavirussen van type B.[2] De circulerende influenzavirussen type A behoren tot de subtypen A(H3N2) en A(H1N1)pdm09 en de influenzavirussen type B zijn antigene varianten behorend tot de Yamagata-lijn of de Victoria-lijn. Griepvirussen van type C veroorzaken in de regel een minder ernstig ziektebeeld en zijn daarom geen target voor vaccinatie en gedetailleerde karakterisering.
Voor het Nationaal Programma Grieppreventie nodigen huisartsen jaarlijks in oktober en november mensen van 60 jaar en ouder en patiënten met een medische indicatie uit om de griepprik te komen halen. Nederland schakelt momenteel over van het gebruik van een trivalent griepvaccin naar een quadrivalent griepvaccin, dat op basis van aanbevelingen van de Wereldgezondheidsorganisatie (WHO) niet alleen de componenten A(H1N1)pdm09 en A(H3N2) bevat maar ook componenten van de beide lijnen van influenzavirus type B.[3] De aanbeveling voor deze vaccinsamenstelling wordt ieder jaar gedaan op grond van epidemiologische en virologische gegevens die binnen het WHO-gecoördineerde internationale surveillancenetwerk worden verkregen. Na de bekendmaking van de aanbevolen vaccinsamenstelling voor het volgende seizoen hebben producenten ongeveer een half jaar de tijd om het vaccin te produceren en distribueren.[4]
Influenzasurveillance
Voor een goede vaccin-geïnduceerde bescherming tegen griep is een goede antigene verwantschap vereist tussen de componenten in het vaccin en de circulerende influenzavirussen van het seizoen. Om dit te bereiken coördineert de WHO het “Global Influenza Surveillance and Response System” (GISRS), dat veranderingen in de circulerende influenzavirussen in kaart brengt.[5] Voor Nederland rapporteert het Nationaal Influenza Centrum wekelijks gegevens over de Nederlandse epidemie aan de WHO en aan het European Centre for Disease Prevention and Control (ECDC). Een selectie van de in Nederland verzamelde en gekarakteriseerde influenzavirussen wordt ook opgestuurd naar het WHO Collaborating Centre in London, dat de Europese data integreert met data van andere continenten, om zo tot een aanbeveling te komen voor het influenzavaccin voor het volgend jaar.
Het Nivel (Nederlands instituut voor onderzoek van de gezondheidszorg) registreert wekelijks het aantal patiënten dat zich met een influenza-achtig ziektebeeld (IAZ) meldt bij de huisarts. Daarnaast wordt het aantal patiënten met een andere acute respiratoire infectie (ARI) geregistreerd. Binnen het Nationaal Influenza Centrum worden door RIVM en Erasmus MC de influenzavirussen verkregen uit luchtwegmonsters van een groot aantal huisarts- en ziekenhuispatiënten met IAZ gekarakteriseerd. Van deze virussen worden de genetische en antigene eigenschappen bepaald en de gevoeligheid voor antivirale middelen vastgesteld. In voorgaande seizoenen werden de genetische eigenschappen van virussen steeds bepaald voor een representatieve subset van de fenotypisch gekarakteriseerde virussen. In het seizoen 2018/2019 werd voor de virussen die waren verkregen uit ziekenhuispatiënten, de volgorde voor het eerst omgedraaid; aan de hand van genetische karakterisering door middel van moderne sequencingtechnieken werd een fenotype voorspeld, dat vervolgens experimenteel werd bevestigd voor een kleine representatieve groep monsters met hoge virusconcentratie, waaruit virus kon worden geïsoleerd.
Resultaten en discussie
De influenza-epidemie van het seizoen 2018/2019
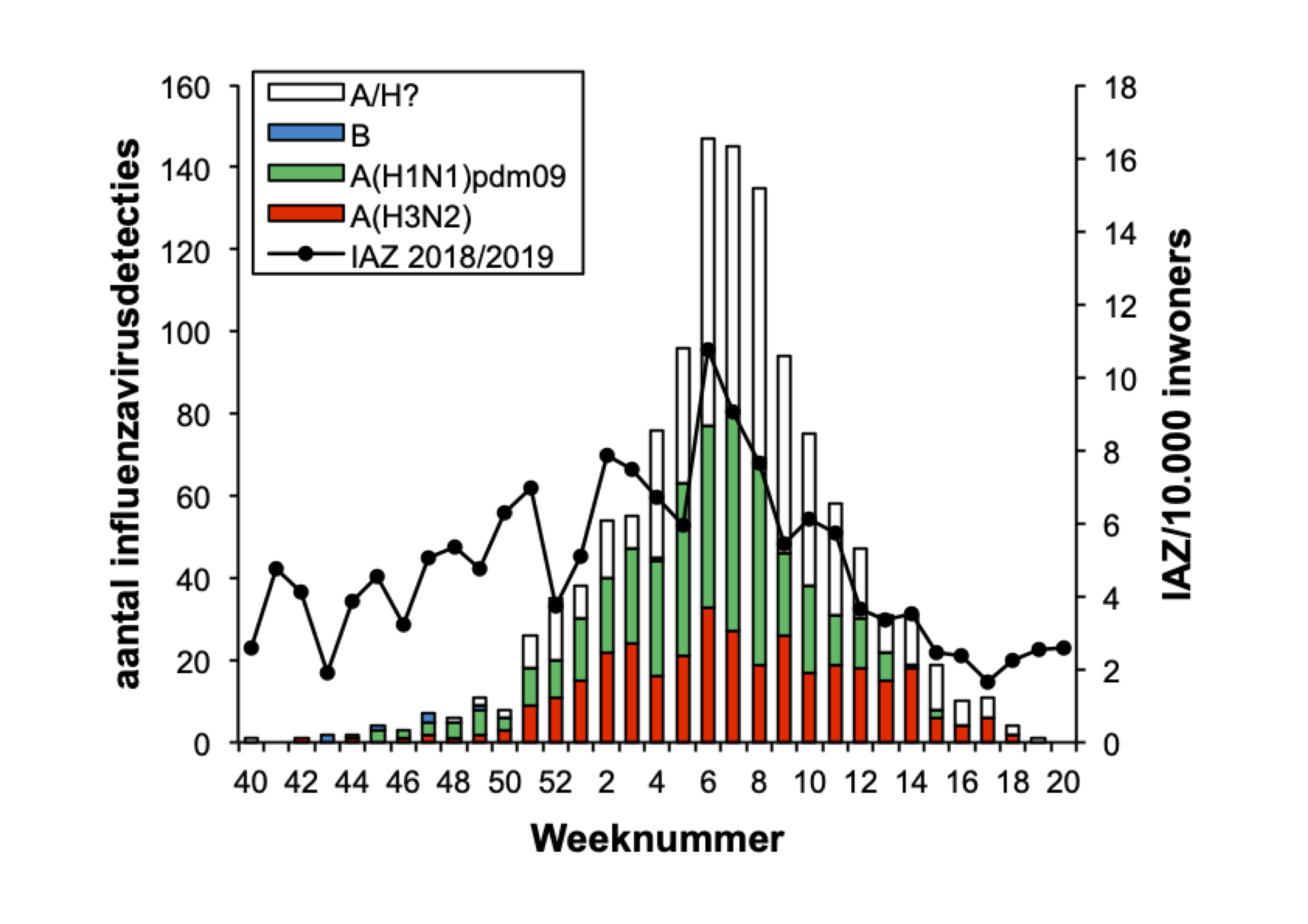
De influenza-epidemie van 2018/2019 duurde in totaal 14 weken, van 10 december 2018 (week 50) tot en met 17 maart 2019 (week 11). In Nederland spreken we van een epidemie wanneer in twee opeenvolgende weken meer dan 5,1 op de 10.000 mensen zich bij hun huisarts melden met IAZ en tegelijkertijd in de neus- en keelmonsters van een deel van deze patiënten influenzavirus wordt aangetoond. De 2018/2019 epidemie duurde dus korter dan de epidemie van vorig jaar (18 weken), maar langer dan het gemiddelde van negen weken van de laatste 25 jaar.[6,7] Dit winterseizoen hadden beduidend minder mensen een IAZ in vergelijking met de drie voorgaande seizoenen (figuur 1). Van de ongebruikelijk hoge sterfte tijdens de griepepidemie van 2017/2018 was dit jaar ook geen sprake. De piek van de epidemie was lager dan in voorgaande jaren, met een maximum van 10,8 IAZ per 10.000 populatie in week 6 (figuur 1). Al met al was er dus sprake van een relatief milde griepepidemie in Nederland.
Overzicht van de onderzochte influenzavirussen
De griepepidemie van 2018/2019 werd gedomineerd door influenzavirussen van type A, waarbij A(H1N1)pdm09- en A(H3N2)-virussen ongeveer even vaak werden gedetecteerd in de peilstations en ziekenhuizen (zie tabel 1). De zeer lage prevalentie van influenzavirus type B was opvallend dit jaar. In de 153 influenzavirus-positieve monsters afgenomen door peilstationhuisartsen bij IAZ-patiënten werd 77 keer (51 procent) een A(H1N1)pdm09-virus gevonden, 74 keer (49 procent) een A(H3N2)-virus en 1 keer (minder dan 1 procent) een influenzavirus type B van de Yamagata-lijn. In de 39 van ARI-patiënten afgenomen influenzavirus-positieve monsters werd 20 keer (51 procent) een A(H3N2)-virus gevonden, 16 keer (41 procent) een A(H1N1)pdm09-virus en 3 keer (8 procent) een influenzavirus type B van de Victoria-lijn (tabel 1 en figuur 2). Van de 1233 door diagnostische ziekenhuislaboratoria aangemelde influenzavirussen waren er 1224 (99 procent) van het A-type en 9 (1 procent) van het B-type. Van de verder gekarakteriseerde A-virussen waren er 387 (53 procent) van het A(H1N1)pdm09-subtype en 339 (47 procent) van het A(H3N2)-subtype. Van de verder gekarakteriseerde B-virussen behoorde er één tot de Yamagata-lijn en één tot de Victoria-lijn (tabel 1 en figuur 3).
Karakterisering van influenza A(H1N1)pdm09-virussen
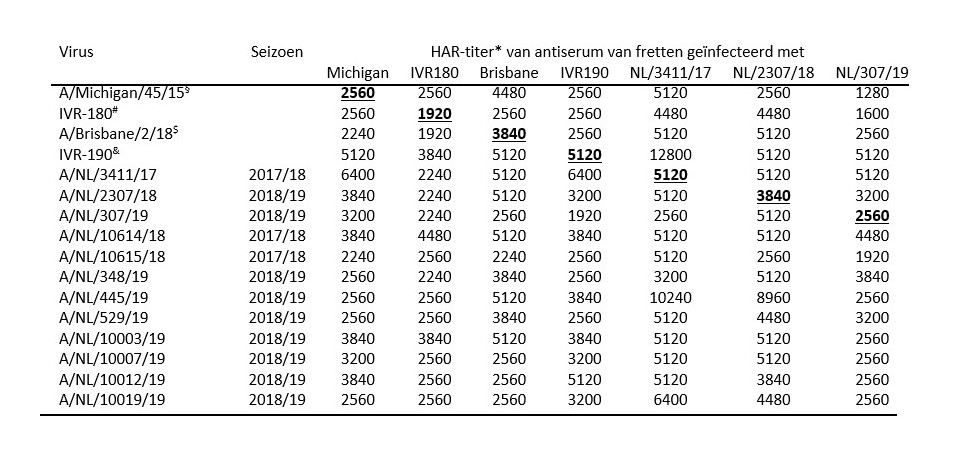
Alle A(H1N1)pdm09-virussen van dit seizoen die werden onderzocht, behoorden genetisch tot clade 6B.1A, met kleine genetische verschillen tussen de virussen onderling (figuur 4). Sinds de pandemie van 2009 hebben de A(H1N1)pdm09-virussen nog weinig antigene variatie laten zien in de hemagglutinatieremmingstest (HAR) met frettensera (minder dan een factor 4 verschil in titers gemeten met sera tegen verschillende A(H1N1)pdm09-stammen). Op basis van humane serologie is voor de vaccinatie voor het seizoen 2017/2018 echter voor het eerst besloten tot een update van deze vaccincomponent naar een A/Michigan/45/2015-achtig virus, die in het seizoen 2018/2019 opnieuw werd aanbevolen. De Nederlandse A(H1N1)pdm09-virussen van het afgelopen seizoen vertoonden goede antigene overeenkomst met de A/Michigan/45/2015-vaccinreferentiestam in de HAR-test met frettensera (tabel 2). De WHO heeft echter opnieuw aanbevolen de A(H1N1)pdm09-vaccincomponent voor het volgend seizoen aan te passen (naar een A/Brisbane/2/2018-achtig virus) op basis van humane serologiedata die verminderde reactiviteit liet zien van sera van kinderen die gevaccineerd werden met de A/Michigan/45/2015-stam tegen een toenemend aantal circulerende virussen.[8]
Karakterisering van influenza A(H3N2)-virussen
De antigene karakterisering van A(H3N2)-virussen is problematisch omdat een groot deel van deze virussen niet of nauwelijks in staat is rode bloedcellen te agglutineren. Dit fenomeen wordt in recente jaren in toenemende mate gezien, en heeft te maken met aminozuursubstituties in het hemagglutinine, waardoor de aviditeit van het virus voor virusreceptoren op onder meer kip- en kalkoenerytrocyten sterk is afgenomen. Wanneer agglutinatie toch wordt waargenomen, is dit vaak het resultaat van een mutatie in het NA-gen, waardoor de agglutinatie via het NA-eiwit kan verlopen.[9,10] Hierdoor kan de HAR-test voor de karakterisering van HA niet of nauwelijks meer worden gebruikt, en moet gewerkt worden met een minder reproduceerbare en meer bewerkelijke virusneutralisatie (VN)-test.[10-12]
De antigene eigenschappen van A(H3N2)-virussen van 2018/2019 waren, evenals vorig seizoen, heterogeen (tabel 3). Zo lieten frettenantisera opgewekt tegen twee recente A(H3N2)-virussen van clades 3C.2a1b en 3C.3a goede homologe neutralisatie zien, maar slechte kruisneutralisatie. Frettensera opgewekt tegen de op eieren geproduceerde A(H3N2)-vaccinstam voor 2018/2019 (NIB-104; A/Singapore/INFIMH-16-0019/2016 clade 3C.2a1) en 2019 voor het zuidelijk halfrond (NIB-112; A/Switzerland/8060/17 clade 3C.2a2) gaven geen of lage kruisreactiviteit tegen de Nederlandse epidemische stammen. Naast de antigene heterogeniteit van de circulerende virussen is dit probleem te wijten aan het feit dat de antigene eigenschappen van recente A(H3N2)-vaccinstammen veranderen na herhaalde passage in eieren, terwijl die eipassage nodig is voor vaccinproductie.
Door de moeizame antigene karakterisering van A(H3N2)-virussen speelt de genetische karakterisering van deze virussen een steeds grotere rol bij de besluitvorming rond de vaccinsamenstelling. Genetische analyse van HA van de in Nederland circulerende A(H3N2)-virussen bevestigde de antigene diversiteit (figuur 4). In Nederland circuleerden afgelopen seizoen virussen behorend tot vooral clade 3C.2a1b (81 procent) en 3C.3a (17 procent) en enkele virussen behorend tot clades 3C.2a1 (n = 1), 3C.2a2 (n = 4), 3C.2a3 (n = 3) en 3C.2a4 (n = 1). In clades 3C.2a1b en 3C.2a3 ontstond nieuwe genetische diversiteit met mogelijk implicaties voor antigene -eigenschappen. De keuze voor de clade 3C.3a-vaccinstam A/Kansas14/17 ter vervanging van de clade 3C.2a1-vaccinstam en de clade 3C.2a2-vaccinstam voor het zuidelijk halfrond is vooral gebaseerd op de toenemende frequentie van detectie van clade 3C.3a-virussen aan het einde van het seizoen 2018/2019 en het gegeven dat de reactiviteit die wordt opgewekt door ei-gekweekte 3C.2a1- en 3C.2a2-vaccins tegen de meeste circulerende A(H3N2)-virussen, beperkt was.[8] Het maken van een gedegen keuze voor een A(H3N2)-vaccincomponent is momenteel vrijwel onmogelijk door de co-circulatie van diverse antigene varianten en de antigene instabiliteit van deze varianten in eieren. Dit was ook de reden dat de WHO in februari 2019 nog geen aanbeveling kon doen voor de vaccinsamenstelling voor het seizoen 2019/2020. Frettensera opgewekt tegen de nieuwe vaccinstam A/Kansas14/17 reageerden wel met recente Nederlandse 3C.3a-virussen, maar niet met 3C.2a1b-virussen (zie tabel 3).
Karakterisering van influenza B-virussen van de Yamagata-lijn
In 2018/2019 werden in Nederland slechts twee influenzavirussen getypeerd als influenzavirus type B van de Yamagata-lijn, waarvan er één werd gekweekt. Beide virussen vielen in dezelfde genetische clade als waartoe de vaccinstam van de Yamagata-lijn B/Phuket/3073/2013 behoort (figuur 4). Het gekweekte virus (B/Nederland/461/2019) liet antigene verschillen zien ten opzichte van de stammen van de voorgaande jaren en de vaccinstam (tabel 4), maar een test met homoloog antiserum moet worden verricht om eenduidige conclusies te trekken over de antigene verwantschap met het vaccin. Volgens het WHO-netwerk is de reactiviteit van de meeste 2018/2019-virussen met sera opgewekt tegen de vaccinstam B/Phuket/3073/2013 nog voldoende en is deze component van het vaccin niet aangepast voor 2019/2020.[8]
Karakterisering van influenza B-virussen van de Victoria-lijn
In het seizoen 2018/2019 werden in Nederland slechts vier influenzavirussen getypeerd als influenzavirus type B van de Victoria-lijn. De twee virussen die genetisch gekarakteriseerd werden, behoorden tot dezelfde genetische clade als waartoe de vaccinstam B/Colorado/6/2017 behoort (figuur 4). Waar de meerderheid van deze virussen een deletie van twee aminozuren heeft ten opzichte van vroegere clades van de Victoria-lijn (posities 162-163), hebben de Nederlandse virussen een deletie van drie aminozuren (posities 162-164). Virussen met een dergelijke deletie van drie aminozuren namen in het seizoen 2018/2019 wereldwijd toe in frequentie. Helaas konden geen van de Nederlandse influenzavirussen van de Victoria-lijn geïsoleerd worden, zodat een analyse van de antigene eigenschappen niet mogelijk was. Uit antigene analyse van vergelijkbare virussen met een deletie van drie aminozuren binnen het WHO-netwerk bleken deze slecht te reageren met antisera opgewekt in fretten tegen de vaccinstam B/Colorado/6/2017 (met een deletie van 2 aminozuren) en B/Brisbane/60/2008 (zonder deleties). Virussen met zowel nul, twee als drie aminozuurdeleties werden goed herkend door humane sera die waren verkregen na vaccinatie tegen B/Colorado/6/2017. Vanwege deze kruisreactiviteit in mensen en de hogere frequentie van detectie van virussen met een deletie van twee aminozuren heeft de WHO besloten de influenzavirus type B-component van de Victoria-lijn niet te veranderen voor het seizoen 2019/2020.[8]
Vaccineffectiviteit
De vaccineffectiviteit gemeten in de huisartsensurveillance tegen in het laboratorium bevestigde infecties met influenzavirus type A en B, inclusief circulerende varianten, was ongeveer 57 procent. De vaccineffectiviteit tegen A(H1N1)pdm09 was ongeveer 60 procent en tegen A(H3N2)-virus 75 procent. In de leeftijdsgroep onder de 60 jaar was de effectiviteit 55 procent en bij mensen boven de 60 was dit 70 procent.[13] Interim-resultaten van het Europese I-Move-netwerk, waar Nederland aan deelneemt, lieten over alle leeftijdsgroepen een iets lagere vaccineffectiviteit zien van 43 procent tegen alle influenzavirussen.[14] Deze vaccineffectiviteit was 71 procent tegen influenza A(H1N1)pdm09-virus, versus -3 procent tegen influenza A(H3N2)-virus. De Nederlandse en Europese schattingen van vaccineffectiviteit komen dus goed overeen voor het A(H1N1)pdm09-subtype, terwijl de schattingen niet goed overeenkomen voor het A(H3N2)-subtype. Er waren geen grote verschillen in circulerende A(H3N2)-virussen in Nederland, vergeleken met de rest van Europa. Het gebrek aan vaccineffectiviteit tegen influenzavirus type A(H3N2), dat in de I-Move-studie vooral bij volwassenen in de werkende leeftijd werd waargenomen, is een reden voor bezorgdheid. Samen met experts uit Canada en de VS, waar vergelijkbare resultaten worden gezien, gaat het I-Move-consortium dit verder evalueren. De vaccineffectiviteit tegen A(H3N2) is daarmee nu al enkele jaren op rij slecht, wat grotendeels te wijten is aan de adaptatie van de A(H3N2)-vaccincomponenten aan de kippeneieren waarin het vaccin wordt geproduceerd en de diversiteit van circulerende virussen.
Vaccinsamenstelling voor het seizoen 2019/2020
Op 21 februari 2019 adviseerde de WHO voor het noordelijk halfrond over de samenstelling van het influenzavaccin, met een update op 21 maart. De update werd noodzakelijk geacht in het licht van de veranderende frequentie van detectie van genetisch en antigeen afwijkende A(H3N2)-virussen in februari. Het uitstel van het advies was uitzonderlijk omdat het productieproces van het griepvaccin ruim een half jaar duurt en de tijdige beschikbaarheid van vaccins voor het seizoen 2019/2020 daardoor in gevaar komt.[4] De uiteindelijk geadviseerde samenstelling was:[8]
- A/Brisbane/02/2018 (H1N1)pdm09-achtig virus;
- A/Kansas/14/2017 (H3N2)-achtig virus;
- B/Colorado/6/2017-achtig virus, van de Victoria-lijn;
- B/Phuket/3073/2013-achtig virus, van de Yamagata-lijn.
Gevoeligheid voor antivirale middelen
Tijdens het seizoen 2018/2019 werden 756 virussen onderzocht op gevoeligheid voor neuraminidaseremmers, waaronder 421 A(H3N2)-, 331 A(H1N1)pdm09- en vier type B-influenzavirussen, en een handvol A(H1N1)pdm09- en A(H3N2)-virussen op gevoeligheid voor M2-blokkers. Drie A(H1N1)pdm09-virussen hadden een sterk verlaagde gevoeligheid voor oseltamivir als gevolg van een 275Y aminozuursubstitutie in het NA-gen. Twee van de patiënten waarin deze virussen gevonden werden, waren opgenomen op de intensive care. Een van deze patiënten was voor afname van het monster behandeld met oseltamivir, voor de ander was de behandelstatus onbekend. De derde patiënt was afkomstig van de huisartspeilstationsurveillance en had geen behandeling met oseltamivir ondergaan voorafgaand aan monsterafname. De A(H3N2)- en A(H1N1)pdm09-influenzavirussen die getest zijn voor resistentie tegen M2-ionkanaalblokkers droegen allemaal de S31N-aminozuursubstitutie die verantwoordelijk is voor resistentie; wereldwijd is dit het geval voor nagenoeg alle A(H3N2)- en A(H1N1)pdm09-virussen.
Conclusie
De influenza-epidemie van 2018/2019 was een relatief milde epidemie, veroorzaakt door hoofdzakelijk A(H3N2)- en A(H1N1)pdm09-virussen. Opvallend was de zeer lage frequentie van detecties van influenza B-virusinfecties (minder dan 1 procent). Het A(H1N2) waarvan wij vorig seizoen melding maakten, is dit jaar niet gevonden in Nederland maar een vergelijkbare (doch niet direct gerelateerde) reassortant werd wel gemeld in Zweden.[15-16] De vaccineffectiviteit lag in 2018/2019 in Nederland rond de 57 procent en bij ouderen zelfs rond de 70 procent, hoewel de schattingen van de effectiviteit door het I-Move-netwerk met name tegen A(H3N2)-virus beduidend lager uitkwamen. Het vinden van een geschikte A(H3N2)-vaccincomponent was ook dit jaar een lastige klus voor het WHO-netwerk, vanwege de co-circulatie van meerdere genetische clades die verschillen vertonen in antigene eigenschappen en die tijdens vaccinproductie in eieren bovendien dusdanig veranderen dat een slechte match ontstaat met epidemische A(H3N2)-virussen.
Dankwoord
Wij zijn zeer erkentelijk voor de bijdragen van de peilstationhuisartsen van Nivel Zorgregistraties eerste lijn (registratie van gevallen van IAZ en verzending van klinische monsters naar het NIC-RIVM) die essentieel zijn voor de influenzasurveillance in Nederland. Zonder de bereidwilligheid van de diagnostische laboratoria om influenzavirusmonsters naar NIC-Erasmus MC te sturen, was deze surveillance evenmin mogelijk geweest.
The authors gratefully acknowledge the generous gift of influenza reference viruses and antisera from Dr. J. McCauley from the World Influenza Centre in London.
De auteurs danken verder R. van Beek, M. Pronk, M. Silva (NIC-Erasmus MC), M. Bagheri, T. Marzec, G. Goderski, S. van den Brink, L. Wijsman, P. Overduin, dr. T. Janssens, drs. D. Reukers, drs. F. Dijkstra en dr. A. Teirlinck (NIC-RIVM) en E. Wentink, M. Hooiveld, Y. Weesie, L. Verberne en J. Gravestein (Nivel) voor de uitstekende technische ondersteuning.