Nederlands Tijdschrift voor Medische Microbiologie Het officiële orgaan van de Nederlandse Vereniging voor Medische Microbiologie (NVMM) informeert lezers over zowel fundamentele als klinische relevante ontwikkelingen binnen het vakgebied. Ook biedt het plaats voor promoties, symposium- en congresverslagen en cursusaankondigingen.
Redactie Dr. Jarne M. van Hattem, Nicolien M. Hanemaaijer, dr. Jaap J. van Hellemond, Maarten Heuvelmans, Jan A. Kaan, dr. Bob Meek, dr. Janette C. Rahamat-Langendoen, Gro L. Vlaspolder, dr. René te Witt
Redactiesecretariaat Alphatekst, Marina Kapteyn Baronie 42 2405 XG Alphen aan den Rijn tel. 06 12076835 marina@alphatekst.nl
Frequentie 4 x per jaar. Alle rechten voorbehouden. Op deze uitgave is het redactiereglement van toepassing. Niets uit deze uitgave mag worden verveelvoudigd, opgeslagen in een geautomatiseerd gegevensbestand of openbaar gemaakt, in enige vorm of op enige wijze, hetzij elektronisch, mechanisch, door fotokopieën, opnamen, of enige andere manier, zonder voorafgaande schriftelijke toestemming van de redactie. De redactie verklaart dat deze uitgave op zorgvuldige wijze en naar beste weten is samengesteld; evenwel kan de redactie op geen enkele wijze instaan voor de juistheid of volledigheid van de informatie. De redactie aanvaardt dan ook geen enkele aansprakelijkheid voor schade, van welke aard ook, die het gevolg is van bedoelde informatie. Gebruikers van deze uitgave wordt met nadruk aangeraden deze informatie niet geïsoleerd te gebruiken, maar af te gaan op hun professionele kennis en ervaring en de te gebruiken informatie te controleren.
<p><span>Thema: Vaccinaties (2)</span></p>
H. de Melker
Editorial
2019;27:nr4
Vaccinaties
Geachte lezer,
Dit nummer van het Nederlands Tijdschrift voor Medische Microbiologie is het tweede themanummer ‘Vaccinaties’.
Vaccinatie is een veld dat continu in beweging is. Nieuwe vaccins (zoals tegen ebola) worden ontwikkeld, plotselinge toename van micro-organisme treden op (meningokokken W) en nieuwe inzichten kunnen aanleiding geven tot aanpassing van vaccinatiebeleid (HPV-vaccinatie ook voor jongens). Veel ziektes zijn door succesvolle vaccinatie (vrijwel) verdwenen of sterk gereduceerd. Wel komen sommige ziekten nog voor doordat vaccinatie geen levenslange bescherming geeft (bof) of door import (mazelen). Ook is er risico op een epidemie door clustering van ongevaccineerden (mazelen, rubella). Ziektelast van andere ziekten blijft hoog door veranderingen in het micro-organisme (influenza, pneumokokken).
Om het vaccinbeleid te optimaliseren is gedegen inzicht nodig in het vóórkomen van de ziekte in combinatie met informatie over (eventuele veranderingen in) het pathogeen. Om surveillancegegevens te kunnen interpreteren is niet alleen epidemiologische en microbiologische kennis nodig, maar ook inzicht in de huidige methoden van diagnostiek in de klinische praktijk. Nieuwe diagnostische methoden, met name op het gebied van de moleculaire diagnostiek en ‘whole genome sequencing’ (WGS), leveren veel relevante informatie. Dit is te lezen in de artikelen uit het huidige themanummer over vaccinatie.
Het artikel van Ron Fouchier en anderen laat zien dat het elk jaar een uitdaging is voor de Wereldgezondheidsorganisatie (WHO) om tot vaccinsamenstelling te komen voor influenza door de telkens veranderde circulatie van influenzavirus. De al lang lopende surveillance door NIVEL en het National Influenza Centrum (Erasmus Rotterdam en RIVM) geeft informatie voor Nederland waarover de WHO wordt geïnformeerd. De auteurs beschrijven de surveillanceresultaten van het influenzaseizoen 2018/2019, waaruit is gebleken dat dit seizoen relatief mild verliep. Uitgebreide aandacht is er voor de karakterisering van influenzavirussen in relatie tot keuze van vaccinsamenstelling en de vaccineffectiviteit die hiermee is bereikt.
Dat inzicht in het virus voor de volksgezondheid belangrijk is, laten ook Rogier Bodewes en medeauteurs zien. Zij schetsen de rol van moleculaire surveillance bij bof, mazelen en rubella waartegen we in Nederland sinds enkele decennia met succes vaccineren. De auteurs lichten toe dat verdere ontwikkeling van sequentietechnieken kan bijdragen aan het vergroten van de moleculaire resolutie waardoor transmissieroutes (import, endemische circulatie) kunnen worden onderzocht en uitbraken kunnen worden gedetecteerd.
Rob Mariman en medeauteurs betogen in hun artikel dat informatie over dragerschap bij zowel jong als oud een belangrijke aanvulling is op ziektesurveillance van invasieve pneumokokkenziekte zoals door het Referentie Laboratorium voor Bacteriële meningitis wordt uitgevoerd. Dragerschap kan worden onderzocht in speeksel van gezonde personen en met behulp van moleculaire diagnostiek en WGS. Dit biedt mogelijk meer zicht op mechanismen die een rol spelen bij de typereplacement die zich voordoet nadat pneumokokkenvaccinatie in 2006 is ingevoerd in het Rijksvaccinatieprogramma.
Bij het artikel van Mirjam Knol en collega’s komt de onvoorspelbare dynamiek van meningokokkenziekte naar voren. Terwijl er een daling optrad van meningokokken B in de jaren 90, waren er forse toenames in invasieve meningokokkenziekte door serogroep C en, meer recent, serogroep W. Dit leidde tot aanpassing van het vaccinatiebeleid. Recent is het meningokokken ACWY-vaccin opgenomen in het Rijksvaccinatieprogramma voor kinderen van 14 maanden en 14 jaar. Het identificeren van de toename en de evaluatie van vaccinatie is mogelijk door al lang lopende surveillance en door het gebruik van nieuwe methoden zoals WGS voor het identificeren van klonale complexen. Ondanks effectieve vaccinatieprogramma’s komt meningokokkenziekte nog steeds voor en is er blijvend aandacht nodig voor vroege herkenning van deze ernstig verlopende ziekte.
Ook in het artikel over Haemophilus influenzae type b (Hib) presenteren Mirjam Knol en Arie van der Ende gegevens over de al lang lopende surveillance van deze invasieve ziekte. Meer dan 25 jaar geleden is gestart met vaccinatie tegen Hib. De ziekte is sterk teruggedrongen, maar de incidentie blijft nog steeds wat hoger dan in andere Europese landen met af en toe tijdelijke toenames in het aantal ziektegevallen. Een oorzaak voor deze toenames werd niet gevonden. De vaccineffectiviteit bleef hoog. Auteurs wijzen verder op een toename van niet-typbeerbare Haemophilus influenzae vooral bij kwetsbare groepen en ouderen.
Joske Hoes beschrijft de introductie van HPV-vaccinatie voor meisjes tien jaar geleden. De lange duur tussen het oplopen van een HPV-infectie en de ontwikkeling van (voorstadia van) kanker betekent dat vroege indicatoren nodig zijn bij de monitoring: het voorkomen van (persisterende) hoogrisico-HPV-infecties. Daarbij wordt gebruikgemaakt van gevoelige genotyperingsmethoden en wordt vaccineffectiviteit aangetoond tegen (persisterende) hoogrisico-HPV-infecties. De impact op (voorstadia van) baarmoederhalskanker zal zichtbaar worden op het moment dat gevaccineerde cohorten de leeftijd van 30 jaar bereiken en in aanmerking komen voor screening op baarmoederhalskanker (2023). Binnen afzienbare tijd wordt het vaccinatieprogramma uitgebreid naar jongensvaccinatie en wordt de mogelijkheid onderzocht van een inhaalvaccinatie tot 26 jaar.
De auteurs van het laatste artikel, Laura Doornekamp et al., beschrijven de status van ebolavaccins en de behandeling ter bestrijding van deze vaak fataal verlopende infectieziekte, waarvan DR Congo al tien uitbraken doormaakte en waardoor ook West-Afrika werd getroffen. Een positieve ontwikkeling is dat de European Medicines Agency (EMA) inmiddels heeft aanbevolen heeft om een voorwaardelijke marktautorisatie te geven voor het rVSV-ZEBOV-GP-vaccin.
Het thema vaccinaties leeft. Dat blijkt ook wel uit het feit dat na dit tweede goedgevulde themanummer Vaccinaties, ook in het volgende nummer van NTMM (maart 2020) onder andere twee vaccingerelateerde onderwerpen aan bod komen, namelijk over rotavirus- en denguevaccinaties.
We attenderen u verder op het ‘In memoriam’ van de zeer gewaardeerde heer J. Hemmes. Het nummer wordt afgesloten met een review van de serologiecursus in mei jl. en de abstracts van de NVMM-najaarsvergadering van 21 november jl.
Hester de Melker, afdelingshoofd Epidemiologie en surveillance van het Rijksvaccinatieprogramma RIVM, gastredacteur, mede namens Gro Vlaspolder en Bert Mulder
<p>J.H. (Jan) Hemmes (1927-2019)</p>
Jan Kaan
In Memoriam
2019;27:nr4
In memoriam
Op 18 oktober 2019 overleed op 92-jarige leeftijd Jan Hemmo Hemmes, in zijn woonplaats Maarssen. Jan Hemmes werd in 1927 geboren in Heteren en groeide op in Utrecht als lid van een gezin met drie kinderen. Zijn vader was arts en later Inspecteur van de Volksgezondheid. Na het gymnasium wilde hij in 1945 weg- en waterbouw studeren. Hij kon in Delft geen kamer vinden en koos toen voor de studie geneeskunde in Utrecht. Na het artsexamen heeft hij zijn militaire dienstplicht vervuld en is vervolgens voor zijn promotie terechtgekomen bij de afdeling Besmettingsleer, onder prof. dr. K.C. Winkler. Hier heeft hij zijn opleiding medische microbiologie afgerond met een promotieonderzoek naar de overleving van virusdeeltjes bij verschillende gradiënten van temperatuur en luchtvochtigheid.[1] De thesis mondde uit in een artikel, “Virus survival as a seasonal factor in influenza and poliomyelitis” [2], dat hij samen met K.C. Winkler en S.M. Kool in 1960 in Nature heeft gepubliceerd. In 1949 leerde hij zijn toekomstige vrouw kennen. Zij trouwden in 1953 en kregen vijf kinderen, één zoon en vier dochters. Omdat zij graag weg wilden uit Utrecht, solliciteerde Jan in 1960 naar een vacature op het Landslaboratorium te Willemstad, Curaçao. Hij werd daar aangenomen en het jonge gezin ondernam de reis naar de Antillen. Uiteindelijk werd Jan hoofd van dit laboratorium. In die periode heeft hij een diag-nostisch medium ontwikkeld, de “7-in”-buis waarmee op grond van zeven reacties (glucose, lactose, saccharose, H2S-productie, urease, beweeglijkheid en indol) een voorspellende identificatie van Salmonella en Shigella gemaakt kan worden. Na zijn terugkeer in 1969, kreeg Jan een aanstelling bij de Streeklaboratoria Pathologie en Microbiologie in Twente en de Gelderse Achterhoek. Hij werkte aanvankelijk samen met bacterioloog A. Marseille, later werd hij benoemd tot adjunct-directeur van de inmiddels in een stichting ondergebrachte laboratoria. Hij heeft de afdelingen Microbiologie altijd in goede samenspraak met zijn collega’s geleid, waarbij het tot consensus komen steeds vooropstond. Ook de samenwerking met de collega’s van de afdeling Pathologie verliep harmonieus. Landelijk heeft Jan vele stenen bijgedragen aan de vooruitgang en uniformering van het vakgebied microbiologie. Daartoe hoorde het op één lijn brengen van de breekpunten. De Werkgroep Richtlijnen Gevoeligheidsbepalingen (WRG) coördineerde de gegevens. Jan voerde in Enschede de agardilutiemethode in. Aflezing van de platen met puntbeënting op groei-geen groei gebeurde al heel vroeg op het laboratorium, waarbij op een terminal de gegevens werden ingevoerd. Deze gegevens werden omgezet naar de (toen nog papieren) uitslag, die dezelfde dag de inzender huisarts en specialist bereikte. Ook konden uitdraaien van verwekkers en spectra worden gemaakt van eerste isolaten voor landelijke vergelijking. In samenwerking met het RIVM (A.J. de Neeling) nam Jan deel aan de landelijke inzameling en aggregatie van resistentiegegevens van zeven streeklaboratoria. Bij zijn vertrek in 1988 werd Jan benoemd tot Officier in de Orde van Oranje-Nassau. Hij ging toen weliswaar met de VUT maar heeft nog lang zijn verknochtheid aan de automatisering niet kunnen weerstaan. In het laboratorium heeft hij nog geruime tijd - voornamelijk epidemiologisch - data-onderzoek verricht. Jan heeft nog vele jaren in goede gezondheid doorgebracht. Toen zijn echtgenote hulpbehoevend werd, zijn zij samen naar een zorginstelling in Maarssen verhuisd, wat voor Jan een moeilijk te nemen stap was. Recent bereikte ons een opnieuw een triest bericht. Op 17 november overleed ook Jans echtgenote Helly van de Lee. Wij wensen de kinderen, klein- en achterkleinkinderen troost toe in het bewustzijn dat Jan Hemmes veel heeft bijgedragen aan de microbiologie in Nederland.
Hemmes JH. De overleving van micro-organismen in de lucht, proefschrift Utrecht 24 maart 1959, Ned Tijdschr Geneeskd 1959;103;36:1820-0.
Hemmes JH, Winkler KC, Kool SM. Virus survival as a seasonal factor in influenza and poliomyelitis. Nature 1960;188;430-1. doi.org/10.1038/188430a0.
<p>Influenzaseizoen 2018/2019 in Nederland<br /><span class="Smaller90">Een relatief milde epidemie met twee subtypen A-virussen</span></p>
Ron Fouchier, Gé Donker, Adam Meijer, Wim van der Hoek, Marit de Lange, Marion Koopmans
Ingezonden
2019;27:nr4
Epidemie, influenza, 2018/2019
Samenvatting De influenza-epidemie van het seizoen 2018/2019 begon in week 50 van 2018 en duurde 14 weken. De epidemie piekte in week 6 van 2019 met een incidentie van 10,8 personen met influenza-achtige ziektebeelden per 10.000 inwoners. Influenza A(H1N1)pdm09- en A(H3N2)-virussen werden ongeveer even vaak gedetecteerd. Opvallend was de zeer sporadische detectie (minder dan 1 procent) van influenzavirus type B dit jaar. Ongeveer 81 procent van de circulerende A(H3N2)-virussen behoorde tot clade 3C.2a1b, waarvoor de antigene overeenkomst met de vaccinstam redelijk tot goed was. Er werd echter, net als elders, een groeiend aandeel (17 procent) clade 3C.3a A(H3N2)-virussen waargenomen, wat een probleem opleverde voor de vaccinkeuze voor volgend jaar. De A(H1N1)pdm09-virussen behoorden genetisch tot clade 6b.1a en vertoonden goede antigene gelijkenis met de vaccinstam. De vaccineffectiviteit tegen laboratoriumbevestigde influenzavirusinfecties was circa 57 procent. Onder de 756 virussen die zijn getest op gevoeligheid voor neuraminidaseremmers waren drie A(H1N1)pdm09-virussen met een sterk verlaagde gevoeligheid voor oseltamivir, in een geval in een onbehandelde patiënt. Alle circulerende influenza A-virussen die werden getest droegen een S31N aminozuursubstitutie in M2 die resistentie tegen ionkanaalblokkers bepaalt. Voor het seizoen 2019/2020 heeft de WHO voor het noordelijk halfrond de volgende vaccinsamenstelling aanbevolen:
Voor A(H1N1)pdm09 een A/Brisbane/02/2018-achtig virus;
Voor A(H3N2) een A/Kansas/14/2017-achtig virus; clade 3C.3a
Voor de B/Victoria-lijn een B/Colorado/6/2017-achtig virus;
Voor de B/Yamagata-lijn een B/Phuket/3073/2013-achtig virus.
Abstract The influenza epidemic of 2018/2019 started in week 50 of 2018 and lasted 14 weeks. The epidemic reached peak incidence in week 6 of 2019, with 10,8 persons presenting influenza-like illness (ILI) per 10.000 inhabitants, marking a relatively mild season. Influenza A(H1N1)pdm09 and A(H3N2 viruses were detected at approximately equivalent frequencies. The sporadic detection (less than 1 per cent) of influenza B viruses was remarkable. Approximately 81 per cent of the circulating A(H3N2) viruses belonged to clade 3C.2a1b, for which the antigenic similarity to the vaccine strain was fair to good. Viruses of clade 3C.3a were detected with increasing frequency nationally (17 per cent) and elsewhere, causing a problem for vaccine strain selection. The A(H1N1)pdm09 viruses belonged to genetic clade 6b.1a and displayed good antigenic similarity to the vaccine strain. The vaccine effectiveness against laboratory-confirmed influenza virus infection was 57 per cent. Among 756 viruses tested for sensitivity to neuraminidase inhibitors, three had strongly reduced sensitivity. All tested influenza A viruses had the S31N amino acid substitution in M2 responsible for resistance to ionchannel inhibitors.
Griep Griep wordt bij mensen veroorzaakt door influenzavirussen van het type A, B of C.[1] Sinds 2009 worden griepepidemieën wereldwijd afwisselend gedomineerd door twee influenzavirussen van type A en twee influenzavirussen van type B.[2] De circulerende influenzavirussen type A behoren tot de subtypen A(H3N2) en A(H1N1)pdm09 en de influenzavirussen type B zijn antigene varianten behorend tot de Yamagata-lijn of de Victoria-lijn. Griepvirussen van type C veroorzaken in de regel een minder ernstig ziektebeeld en zijn daarom geen target voor vaccinatie en gedetailleerde karakterisering.
Voor het Nationaal Programma Grieppreventie nodigen huisartsen jaarlijks in oktober en november mensen van 60 jaar en ouder en patiënten met een medische indicatie uit om de griepprik te komen halen. Nederland schakelt momenteel over van het gebruik van een trivalent griepvaccin naar een quadrivalent griepvaccin, dat op basis van aanbevelingen van de Wereldgezondheidsorganisatie (WHO) niet alleen de componenten A(H1N1)pdm09 en A(H3N2) bevat maar ook componenten van de beide lijnen van influenzavirus type B.[3] De aanbeveling voor deze vaccinsamenstelling wordt ieder jaar gedaan op grond van epidemiologische en virologische gegevens die binnen het WHO-gecoördineerde internationale surveillancenetwerk worden verkregen. Na de bekendmaking van de aanbevolen vaccinsamenstelling voor het volgende seizoen hebben producenten ongeveer een half jaar de tijd om het vaccin te produceren en distribueren.[4]
Influenzasurveillance Voor een goede vaccin-geïnduceerde bescherming tegen griep is een goede antigene verwantschap vereist tussen de componenten in het vaccin en de circulerende influenzavirussen van het seizoen. Om dit te bereiken coördineert de WHO het “Global Influenza Surveillance and Response System” (GISRS), dat veranderingen in de circulerende influenzavirussen in kaart brengt.[5] Voor Nederland rapporteert het Nationaal Influenza Centrum wekelijks gegevens over de Nederlandse epidemie aan de WHO en aan het European Centre for Disease Prevention and Control (ECDC). Een selectie van de in Nederland verzamelde en gekarakteriseerde influenzavirussen wordt ook opgestuurd naar het WHO Collaborating Centre in London, dat de Europese data integreert met data van andere continenten, om zo tot een aanbeveling te komen voor het influenzavaccin voor het volgend jaar.
Het Nivel (Nederlands instituut voor onderzoek van de gezondheidszorg) registreert wekelijks het aantal patiënten dat zich met een influenza-achtig ziektebeeld (IAZ) meldt bij de huisarts. Daarnaast wordt het aantal patiënten met een andere acute respiratoire infectie (ARI) geregistreerd. Binnen het Nationaal Influenza Centrum worden door RIVM en Erasmus MC de influenzavirussen verkregen uit luchtwegmonsters van een groot aantal huisarts- en ziekenhuispatiënten met IAZ gekarakteriseerd. Van deze virussen worden de genetische en antigene eigenschappen bepaald en de gevoeligheid voor antivirale middelen vastgesteld. In voorgaande seizoenen werden de genetische eigenschappen van virussen steeds bepaald voor een representatieve subset van de fenotypisch gekarakteriseerde virussen. In het seizoen 2018/2019 werd voor de virussen die waren verkregen uit ziekenhuispatiënten, de volgorde voor het eerst omgedraaid; aan de hand van genetische karakterisering door middel van moderne sequencingtechnieken werd een fenotype voorspeld, dat vervolgens experimenteel werd bevestigd voor een kleine representatieve groep monsters met hoge virusconcentratie, waaruit virus kon worden geïsoleerd.
Resultaten en discussie
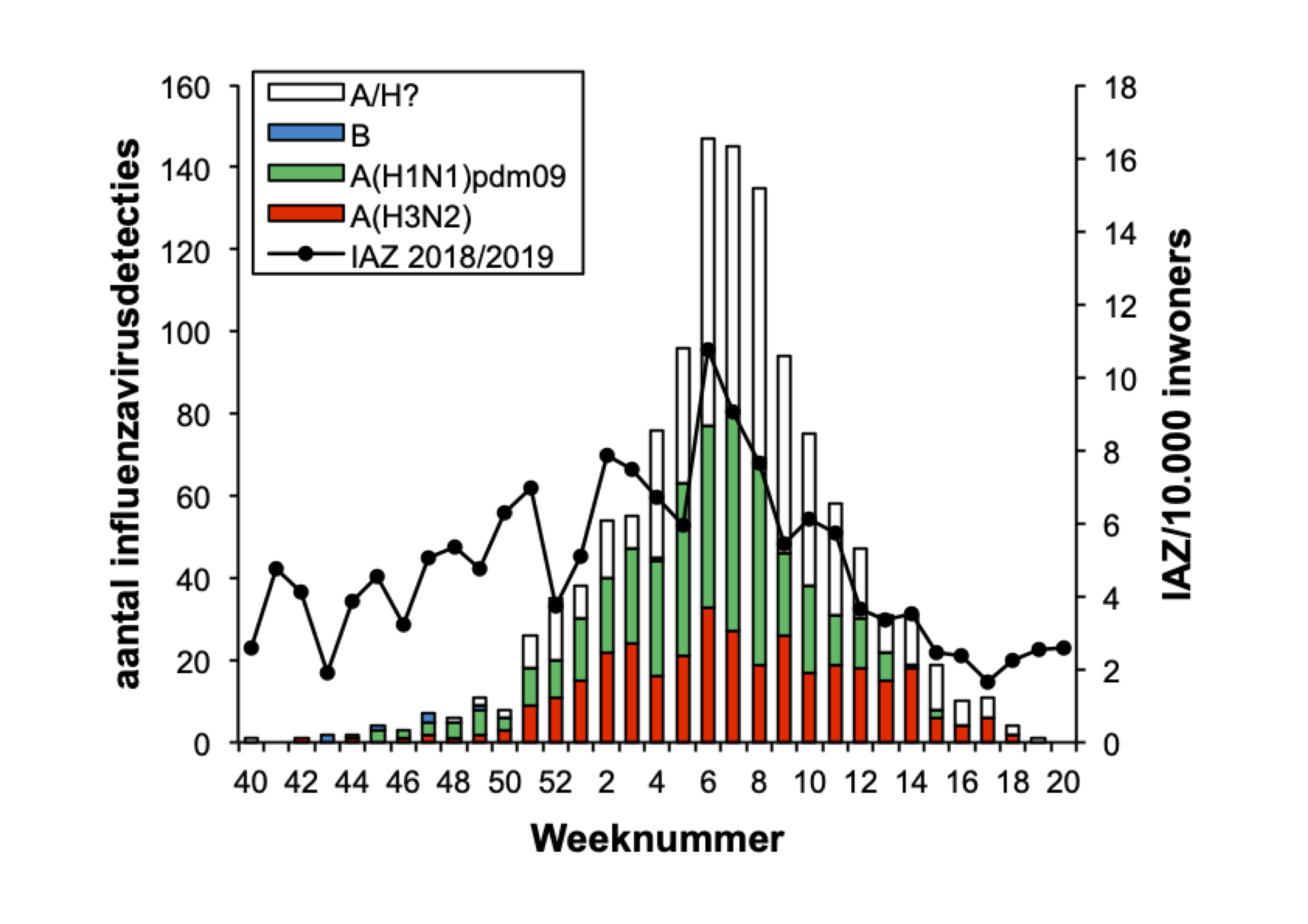
De influenza-epidemie van het seizoen 2018/2019 De influenza-epidemie van 2018/2019 duurde in totaal 14 weken, van 10 december 2018 (week 50) tot en met 17 maart 2019 (week 11). In Nederland spreken we van een epidemie wanneer in twee opeenvolgende weken meer dan 5,1 op de 10.000 mensen zich bij hun huisarts melden met IAZ en tegelijkertijd in de neus- en keelmonsters van een deel van deze patiënten influenzavirus wordt aangetoond. De 2018/2019 epidemie duurde dus korter dan de epidemie van vorig jaar (18 weken), maar langer dan het gemiddelde van negen weken van de laatste 25 jaar.[6,7] Dit winterseizoen hadden beduidend minder mensen een IAZ in vergelijking met de drie voorgaande seizoenen (figuur 1). Van de ongebruikelijk hoge sterfte tijdens de griepepidemie van 2017/2018 was dit jaar ook geen sprake. De piek van de epidemie was lager dan in voorgaande jaren, met een maximum van 10,8 IAZ per 10.000 populatie in week 6 (figuur 1). Al met al was er dus sprake van een relatief milde griepepidemie in Nederland.
Overzicht van de onderzochte influenzavirussen De griepepidemie van 2018/2019 werd gedomineerd door influenzavirussen van type A, waarbij A(H1N1)pdm09- en A(H3N2)-virussen ongeveer even vaak werden gedetecteerd in de peilstations en ziekenhuizen (zie tabel 1). De zeer lage prevalentie van influenzavirus type B was opvallend dit jaar. In de 153 influenzavirus-positieve monsters afgenomen door peilstationhuisartsen bij IAZ-patiënten werd 77 keer (51 procent) een A(H1N1)pdm09-virus gevonden, 74 keer (49 procent) een A(H3N2)-virus en 1 keer (minder dan 1 procent) een influenzavirus type B van de Yamagata-lijn. In de 39 van ARI-patiënten afgenomen influenzavirus-positieve monsters werd 20 keer (51 procent) een A(H3N2)-virus gevonden, 16 keer (41 procent) een A(H1N1)pdm09-virus en 3 keer (8 procent) een influenzavirus type B van de Victoria-lijn (tabel 1 en figuur 2). Van de 1233 door diagnostische ziekenhuislaboratoria aangemelde influenzavirussen waren er 1224 (99 procent) van het A-type en 9 (1 procent) van het B-type. Van de verder gekarakteriseerde A-virussen waren er 387 (53 procent) van het A(H1N1)pdm09-subtype en 339 (47 procent) van het A(H3N2)-subtype. Van de verder gekarakteriseerde B-virussen behoorde er één tot de Yamagata-lijn en één tot de Victoria-lijn (tabel 1 en figuur 3).
Karakterisering van influenza A(H1N1)pdm09-virussen Alle A(H1N1)pdm09-virussen van dit seizoen die werden onderzocht, behoorden genetisch tot clade 6B.1A, met kleine genetische verschillen tussen de virussen onderling (figuur 4). Sinds de pandemie van 2009 hebben de A(H1N1)pdm09-virussen nog weinig antigene variatie laten zien in de hemagglutinatieremmingstest (HAR) met frettensera (minder dan een factor 4 verschil in titers gemeten met sera tegen verschillende A(H1N1)pdm09-stammen). Op basis van humane serologie is voor de vaccinatie voor het seizoen 2017/2018 echter voor het eerst besloten tot een update van deze vaccincomponent naar een A/Michigan/45/2015-achtig virus, die in het seizoen 2018/2019 opnieuw werd aanbevolen. De Nederlandse A(H1N1)pdm09-virussen van het afgelopen seizoen vertoonden goede antigene overeenkomst met de A/Michigan/45/2015-vaccinreferentiestam in de HAR-test met frettensera (tabel 2). De WHO heeft echter opnieuw aanbevolen de A(H1N1)pdm09-vaccincomponent voor het volgend seizoen aan te passen (naar een A/Brisbane/2/2018-achtig virus) op basis van humane serologiedata die verminderde reactiviteit liet zien van sera van kinderen die gevaccineerd werden met de A/Michigan/45/2015-stam tegen een toenemend aantal circulerende virussen.[8]
Karakterisering van influenza A(H3N2)-virussen De antigene karakterisering van A(H3N2)-virussen is problematisch omdat een groot deel van deze virussen niet of nauwelijks in staat is rode bloedcellen te agglutineren. Dit fenomeen wordt in recente jaren in toenemende mate gezien, en heeft te maken met aminozuursubstituties in het hemagglutinine, waardoor de aviditeit van het virus voor virusreceptoren op onder meer kip- en kalkoenerytrocyten sterk is afgenomen. Wanneer agglutinatie toch wordt waargenomen, is dit vaak het resultaat van een mutatie in het NA-gen, waardoor de agglutinatie via het NA-eiwit kan verlopen.[9,10] Hierdoor kan de HAR-test voor de karakterisering van HA niet of nauwelijks meer worden gebruikt, en moet gewerkt worden met een minder reproduceerbare en meer bewerkelijke virusneutralisatie (VN)-test.[10-12]
De antigene eigenschappen van A(H3N2)-virussen van 2018/2019 waren, evenals vorig seizoen, heterogeen (tabel 3). Zo lieten frettenantisera opgewekt tegen twee recente A(H3N2)-virussen van clades 3C.2a1b en 3C.3a goede homologe neutralisatie zien, maar slechte kruisneutralisatie. Frettensera opgewekt tegen de op eieren geproduceerde A(H3N2)-vaccinstam voor 2018/2019 (NIB-104; A/Singapore/INFIMH-16-0019/2016 clade 3C.2a1) en 2019 voor het zuidelijk halfrond (NIB-112; A/Switzerland/8060/17 clade 3C.2a2) gaven geen of lage kruisreactiviteit tegen de Nederlandse epidemische stammen. Naast de antigene heterogeniteit van de circulerende virussen is dit probleem te wijten aan het feit dat de antigene eigenschappen van recente A(H3N2)-vaccinstammen veranderen na herhaalde passage in eieren, terwijl die eipassage nodig is voor vaccinproductie. Door de moeizame antigene karakterisering van A(H3N2)-virussen speelt de genetische karakterisering van deze virussen een steeds grotere rol bij de besluitvorming rond de vaccinsamenstelling. Genetische analyse van HA van de in Nederland circulerende A(H3N2)-virussen bevestigde de antigene diversiteit (figuur 4). In Nederland circuleerden afgelopen seizoen virussen behorend tot vooral clade 3C.2a1b (81 procent) en 3C.3a (17 procent) en enkele virussen behorend tot clades 3C.2a1 (n = 1), 3C.2a2 (n = 4), 3C.2a3 (n = 3) en 3C.2a4 (n = 1). In clades 3C.2a1b en 3C.2a3 ontstond nieuwe genetische diversiteit met mogelijk implicaties voor antigene -eigenschappen. De keuze voor de clade 3C.3a-vaccinstam A/Kansas14/17 ter vervanging van de clade 3C.2a1-vaccinstam en de clade 3C.2a2-vaccinstam voor het zuidelijk halfrond is vooral gebaseerd op de toenemende frequentie van detectie van clade 3C.3a-virussen aan het einde van het seizoen 2018/2019 en het gegeven dat de reactiviteit die wordt opgewekt door ei-gekweekte 3C.2a1- en 3C.2a2-vaccins tegen de meeste circulerende A(H3N2)-virussen, beperkt was.[8] Het maken van een gedegen keuze voor een A(H3N2)-vaccincomponent is momenteel vrijwel onmogelijk door de co-circulatie van diverse antigene varianten en de antigene instabiliteit van deze varianten in eieren. Dit was ook de reden dat de WHO in februari 2019 nog geen aanbeveling kon doen voor de vaccinsamenstelling voor het seizoen 2019/2020. Frettensera opgewekt tegen de nieuwe vaccinstam A/Kansas14/17 reageerden wel met recente Nederlandse 3C.3a-virussen, maar niet met 3C.2a1b-virussen (zie tabel 3).
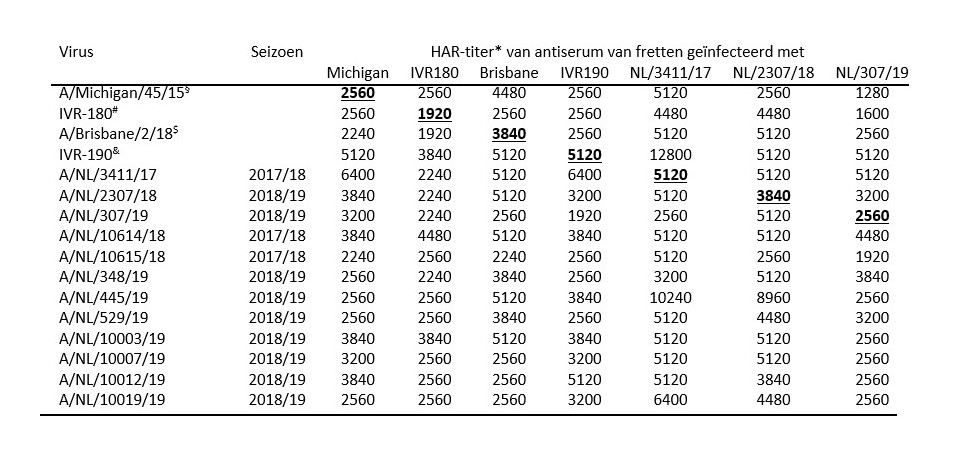
Karakterisering van influenza B-virussen van de Yamagata-lijn In 2018/2019 werden in Nederland slechts twee influenzavirussen getypeerd als influenzavirus type B van de Yamagata-lijn, waarvan er één werd gekweekt. Beide virussen vielen in dezelfde genetische clade als waartoe de vaccinstam van de Yamagata-lijn B/Phuket/3073/2013 behoort (figuur 4). Het gekweekte virus (B/Nederland/461/2019) liet antigene verschillen zien ten opzichte van de stammen van de voorgaande jaren en de vaccinstam (tabel 4), maar een test met homoloog antiserum moet worden verricht om eenduidige conclusies te trekken over de antigene verwantschap met het vaccin. Volgens het WHO-netwerk is de reactiviteit van de meeste 2018/2019-virussen met sera opgewekt tegen de vaccinstam B/Phuket/3073/2013 nog voldoende en is deze component van het vaccin niet aangepast voor 2019/2020.[8]
Karakterisering van influenza B-virussen van de Victoria-lijn In het seizoen 2018/2019 werden in Nederland slechts vier influenzavirussen getypeerd als influenzavirus type B van de Victoria-lijn. De twee virussen die genetisch gekarakteriseerd werden, behoorden tot dezelfde genetische clade als waartoe de vaccinstam B/Colorado/6/2017 behoort (figuur 4). Waar de meerderheid van deze virussen een deletie van twee aminozuren heeft ten opzichte van vroegere clades van de Victoria-lijn (posities 162-163), hebben de Nederlandse virussen een deletie van drie aminozuren (posities 162-164). Virussen met een dergelijke deletie van drie aminozuren namen in het seizoen 2018/2019 wereldwijd toe in frequentie. Helaas konden geen van de Nederlandse influenzavirussen van de Victoria-lijn geïsoleerd worden, zodat een analyse van de antigene eigenschappen niet mogelijk was. Uit antigene analyse van vergelijkbare virussen met een deletie van drie aminozuren binnen het WHO-netwerk bleken deze slecht te reageren met antisera opgewekt in fretten tegen de vaccinstam B/Colorado/6/2017 (met een deletie van 2 aminozuren) en B/Brisbane/60/2008 (zonder deleties). Virussen met zowel nul, twee als drie aminozuurdeleties werden goed herkend door humane sera die waren verkregen na vaccinatie tegen B/Colorado/6/2017. Vanwege deze kruisreactiviteit in mensen en de hogere frequentie van detectie van virussen met een deletie van twee aminozuren heeft de WHO besloten de influenzavirus type B-component van de Victoria-lijn niet te veranderen voor het seizoen 2019/2020.[8]
Vaccineffectiviteit De vaccineffectiviteit gemeten in de huisartsensurveillance tegen in het laboratorium bevestigde infecties met influenzavirus type A en B, inclusief circulerende varianten, was ongeveer 57 procent. De vaccineffectiviteit tegen A(H1N1)pdm09 was ongeveer 60 procent en tegen A(H3N2)-virus 75 procent. In de leeftijdsgroep onder de 60 jaar was de effectiviteit 55 procent en bij mensen boven de 60 was dit 70 procent.[13] Interim-resultaten van het Europese I-Move-netwerk, waar Nederland aan deelneemt, lieten over alle leeftijdsgroepen een iets lagere vaccineffectiviteit zien van 43 procent tegen alle influenzavirussen.[14] Deze vaccineffectiviteit was 71 procent tegen influenza A(H1N1)pdm09-virus, versus -3 procent tegen influenza A(H3N2)-virus. De Nederlandse en Europese schattingen van vaccineffectiviteit komen dus goed overeen voor het A(H1N1)pdm09-subtype, terwijl de schattingen niet goed overeenkomen voor het A(H3N2)-subtype. Er waren geen grote verschillen in circulerende A(H3N2)-virussen in Nederland, vergeleken met de rest van Europa. Het gebrek aan vaccineffectiviteit tegen influenzavirus type A(H3N2), dat in de I-Move-studie vooral bij volwassenen in de werkende leeftijd werd waargenomen, is een reden voor bezorgdheid. Samen met experts uit Canada en de VS, waar vergelijkbare resultaten worden gezien, gaat het I-Move-consortium dit verder evalueren. De vaccineffectiviteit tegen A(H3N2) is daarmee nu al enkele jaren op rij slecht, wat grotendeels te wijten is aan de adaptatie van de A(H3N2)-vaccincomponenten aan de kippeneieren waarin het vaccin wordt geproduceerd en de diversiteit van circulerende virussen.
Vaccinsamenstelling voor het seizoen 2019/2020 Op 21 februari 2019 adviseerde de WHO voor het noordelijk halfrond over de samenstelling van het influenzavaccin, met een update op 21 maart. De update werd noodzakelijk geacht in het licht van de veranderende frequentie van detectie van genetisch en antigeen afwijkende A(H3N2)-virussen in februari. Het uitstel van het advies was uitzonderlijk omdat het productieproces van het griepvaccin ruim een half jaar duurt en de tijdige beschikbaarheid van vaccins voor het seizoen 2019/2020 daardoor in gevaar komt.[4] De uiteindelijk geadviseerde samenstelling was:[8]
A/Brisbane/02/2018 (H1N1)pdm09-achtig virus;
A/Kansas/14/2017 (H3N2)-achtig virus;
B/Colorado/6/2017-achtig virus, van de Victoria-lijn;
B/Phuket/3073/2013-achtig virus, van de Yamagata-lijn.
Gevoeligheid voor antivirale middelen Tijdens het seizoen 2018/2019 werden 756 virussen onderzocht op gevoeligheid voor neuraminidaseremmers, waaronder 421 A(H3N2)-, 331 A(H1N1)pdm09- en vier type B-influenzavirussen, en een handvol A(H1N1)pdm09- en A(H3N2)-virussen op gevoeligheid voor M2-blokkers. Drie A(H1N1)pdm09-virussen hadden een sterk verlaagde gevoeligheid voor oseltamivir als gevolg van een 275Y aminozuursubstitutie in het NA-gen. Twee van de patiënten waarin deze virussen gevonden werden, waren opgenomen op de intensive care. Een van deze patiënten was voor afname van het monster behandeld met oseltamivir, voor de ander was de behandelstatus onbekend. De derde patiënt was afkomstig van de huisartspeilstationsurveillance en had geen behandeling met oseltamivir ondergaan voorafgaand aan monsterafname. De A(H3N2)- en A(H1N1)pdm09-influenzavirussen die getest zijn voor resistentie tegen M2-ionkanaalblokkers droegen allemaal de S31N-aminozuursubstitutie die verantwoordelijk is voor resistentie; wereldwijd is dit het geval voor nagenoeg alle A(H3N2)- en A(H1N1)pdm09-virussen.
Conclusie De influenza-epidemie van 2018/2019 was een relatief milde epidemie, veroorzaakt door hoofdzakelijk A(H3N2)- en A(H1N1)pdm09-virussen. Opvallend was de zeer lage frequentie van detecties van influenza B-virusinfecties (minder dan 1 procent). Het A(H1N2) waarvan wij vorig seizoen melding maakten, is dit jaar niet gevonden in Nederland maar een vergelijkbare (doch niet direct gerelateerde) reassortant werd wel gemeld in Zweden.[15-16] De vaccineffectiviteit lag in 2018/2019 in Nederland rond de 57 procent en bij ouderen zelfs rond de 70 procent, hoewel de schattingen van de effectiviteit door het I-Move-netwerk met name tegen A(H3N2)-virus beduidend lager uitkwamen. Het vinden van een geschikte A(H3N2)-vaccincomponent was ook dit jaar een lastige klus voor het WHO-netwerk, vanwege de co-circulatie van meerdere genetische clades die verschillen vertonen in antigene eigenschappen en die tijdens vaccinproductie in eieren bovendien dusdanig veranderen dat een slechte match ontstaat met epidemische A(H3N2)-virussen.
Dankwoord Wij zijn zeer erkentelijk voor de bijdragen van de peilstationhuisartsen van Nivel Zorgregistraties eerste lijn (registratie van gevallen van IAZ en verzending van klinische monsters naar het NIC-RIVM) die essentieel zijn voor de influenzasurveillance in Nederland. Zonder de bereidwilligheid van de diagnostische laboratoria om influenzavirusmonsters naar NIC-Erasmus MC te sturen, was deze surveillance evenmin mogelijk geweest.
The authors gratefully acknowledge the generous gift of influenza reference viruses and antisera from Dr. J. McCauley from the World Influenza Centre in London.
De auteurs danken verder R. van Beek, M. Pronk, M. Silva (NIC-Erasmus MC), M. Bagheri, T. Marzec, G. Goderski, S. van den Brink, L. Wijsman, P. Overduin, dr. T. Janssens, drs. D. Reukers, drs. F. Dijkstra en dr. A. Teirlinck (NIC-RIVM) en E. Wentink, M. Hooiveld, Y. Weesie, L. Verberne en J. Gravestein (Nivel) voor de uitstekende technische ondersteuning.
Lindstrom SE, Hiromoto Y, Nishimura H, et al. Comparative analysis of evolutionary mechanisms of the hemagglutinin and three internal protein genes of influenza B virus: multiple cocirculating lineages and frequent reassortment of the NP, M, and NS genes. J Virol. 1999;73:4413-26.
WHO. Recommended composition of influenza virus vaccines for use in the 2018–2019 northern hemisphere influenza season. Wkly Epidemiol Rec. 2018;93:133-41.
Meijer A, Timmermans J, Donker GA, et al. Elk jaar een nieuw griepvaccin. Hoe wordt de samenstelling ervan bepaald? Tijdschr Infect. 2017;12:33-40.
Hay AJ, McCauley JW. The WHO global influenza surveillance and response system (GISRS)-A future perspective. Influenza Other Respir Viruses. 2018;12:551-7.
Fouchier RA, Meijer A, Donker GA, et al. Influenzaseizoen 2017/2018 in Nederland. Ned Tijdschr Med Microbiol. 2019;27:52-63.
Mögling R, Richard MJ, van der Vliet S, et al. Neuraminidase- mediated hemagglutination of recent human influenza A(H3N2) viruses is determined by arginine 150 flanking the neuraminidase catalytic site. J Gen Virol 2017;98:1274-81.
Lin YP, Gregory V, Collins P, et al. Neuraminidase receptor-binding variants of human influenza A(H3N2) viruses resulting from substitution of aspartic acid 151 in the catalytic site: a role in virus attachment? J Virol. 2010;84:6769-81.
Van Baalen CA, Els C, Sprong L, et al. Detection of non-hemagglutinating influenza A(H3) viruses by ELISA in quantitative influenza virus culture. J Clin Microbiol. 2014;52:1672-7.
Van Baalen CA, Jeeninga R, Penders G, et al. ViroSpotTM microneutralization assay for antigenic characterization of influenza viruses. Vaccine. 2017;35:46-52.
Reukers DFM, van Asten L, Brandsema PS, et al. Annual Report Surveillance of influenza and other respiratory infections in the Netherlands: winter 2018/2019. RIVM report no. 2019-0079.
Kissling E, Rose A, Emborg HD, et al. Interim 2018/19 influenza vaccine effectiveness: six European studies, October 2018 to January 2019. Euro Surveill. 2019;24(8).
Meijer A, Swaan CM, Voerknecht M, et al. Case of seasonal reassortant A(H1N2) influenza virus infection, the Netherlands, March 2018. Euro Surveill. 2018;23:pii=18-00160.
Wiman Å, Enkirch T, Carnahan A, et al. Novel influenza A(H1N2) seasonal reassortant identified in a patient sample, Sweden, January 2019. Euro Surveill. 2019;24(9).
Erasmus MC, afdeling Viroscience, Nationaal Influenza Centrum, Rotterdam, prof. dr. R.A.M. Fouchier, prof. dr. M.P.G. Koopmans, virologen. Nivel Zorgregistraties eerste lijn-peilstations, Utrecht, dr. G.A. Donker, huisarts-epidemioloog. Rijksinstituut voor Volksgezondheid en Milieu (RIVM, Centrum Infectieziektebestrijding, Nationaal Influenza Centrum, Bilthoven, dr. A. Meijer, viroloog, dr. W. van der Hoek, arts- epidemioloog, drs. M.M.A. de Lange, epidemioloog. Correspondentieadres: prof. dr. R.A.M. Fouchier (r.fouchier@erasmusmc.nl).
<p><span class="Smaller90"><strong>Figuur 1.</strong> Klinische influenza-activiteit in Nederland in de seizoenen 2015/2016 tot en met 2018/2019, weergegeven als het wekelijkse aantal patiënten met een influenza-achtig ziektebeeld (IAZ) per 10.000 inwoners, aangemeld bij de huisartsenpeilstations van week 40 tot en met week 20 van het volgende jaar. Bron: Nivel Zorgregistraties eerste lijn. Week 53 geldt alleen voor seizoen 2015/2016.</span></p>
<p><span class="Smaller90"><strong>Figuur 2.</strong> Virusdetecties in het influenzaseizoen 2018/2019 in de door peilstations afgenomen monsters van patiënten gediagnostiseerd met een influenza-achtig ziektebeeld (IAZ) (A), of een andere acute respiratoire infectie (ARI) (B). Afgebeeld zijn de wekelijkse aantallen detecties van influenzavirus, opgesplitst naar (sub)type. De aantallen zijn weergegeven als balken en als percentage van de monsters waarin een influenzavirus werd aangetroffen (stippellijn), af te lezen op de linker verticale as. Ook het aantal IAZ per 10.000 inwoners per week is weergegeven, af te lezen op de rechter verticale as.</span></p>
A.
B.
<p><span class="Smaller90"><strong>Tabel 1.</strong> Detectie van influenzavirussen in luchtwegmonsters van patiënten met IAZ en ARI die ingestuurd werden vanuit de huisartsenpeilstations en vanuit diagnostische laboratoria van ziekenhuizen van week 40 van 2018 tot en met week 20 van 2019.</span></p>
Aantallen influenzavirusdetecties (%)*
Type A
Type B
Bron van virusdetecties§
H3N2
H1N1pdm09
Anders#
Victoria
Yamagata
Anders#
IAZ-patiënten
74 (49%)
77 (51%)
1
0 (0%)
1 (< 1%)
0
ARI-patiënten
20 (51%)
16 (41%)
0
3 (8%)
0 (0%)
0
Ziekenhuizen
339 (47%)
387 (53%)
498
1 (< 1%)
1 (< 1%)
7
<p><span class="Smaller90"><strong>Figuur 3.</strong> Virusdetecties in het influenzaseizoen 2018/2019 in monsters ingestuurd vanuit diagnostische ziekenhuislaboratoria. Afgebeeld zijn de wekelijkse aantallen detecties van influenzavirus, opgesplitst naar (sub)type. De aantallen zijn weergegeven als balken, af te lezen op de linker verticale as. Ook het aantal IAZ per 10.000 inwoners per week is weergegeven, af te lezen op de rechter verticale as. </span></p>
<p><span class="Smaller90"><strong>Tabel 2.</strong> Antigene karakterisering van Nederlandse influenza A(H1N1)pdm09-virussen uit het seizoen 2018/2019.</span></p>
<p><span class="Smaller90"><strong>Tabel 3.</strong> Antigene karakterisering van Nederlandse influenza A(H3N2)-virussen uit het seizoen 2018/2019.</span></p>
Virus
Clade
Titer* van antiserum van fretten geïnfecteerd met
NIB-104§
NIB-112#
NL/314/19
NL/384/19
X-327$
3C.2a1
3C.2a2
3C.2a1b
3C.3a
3C.3a
NIB-104§
3C.2a1
640
2560
320
< 20
320
Switzerland/8060/17
3C.2a2
80
80
40
< 20
< 20
NL/314/19
3C.2a1b
20
< 20
640
< 20
< 20
NL/371/19
3C.2a1b
< 20
< 20
160
< 20
< 20
NL/10009/19
3C.2a1b
20
40
640
< 20
< 20
NL/384/19
3C.3a
< 20
< 20
< 20
160
40
NL/10002/19
3C.3a
< 20
< 20
< 20
< 20
80
NL/10006/19
3C.3a
< 20
< 20
< 20
320
80
<p><span class="Smaller90"><strong>Tabel 4.</strong> Antigene karakterisering van Nederlands influenza B virus van de Yamagata-lijn uit het seizoen 2018/2019.</span></p>
Virus
Seizoen
HAR-titer* van antiserum van fretten geïnfecteerd met
Wisconsin
Phuket
NL/3066/15
NL/2424/17
NL/4136/17
B/Wisconsin/1/10§
1280
1280
480
480
160
B/Phuket/3073/13#
1920
1280
120
160
80
B/NL/3066/15
2015/16
400
400
1120
960
960
B/NL/2424/17
2016/17
160
280
640
1120
320
B/NL/4136/17
2017/18
640
320
1280
1280
800
B/NL/461/19
2018/19
80
40
80
320
70
<p><span class="Smaller90"><strong>Figuur 4.</strong> Fylogenetische analyse van de influenzavirussen die circuleerden in Nederland in het seizoen 2018/2019. De fylogenetische bomen voor (A) influenzavirus A(H1N1)pdm09, (B) influenzavirus A(H3N2), (C) influenza B-virus van de Yamagata-lijn en (D) influenza B-virus van de Victoria-lijn zijn berekend met de hemagglutinine aminozuursequenties zonder signaalpeptide en stopcodon. De bomen zijn gemaakt met Molecular Evolutionary Genetics Analysis (MEGA X) versie 10.0.5, waarin de Maximum Likelihood methode is gebruikt met als subsitutiemodel ‘JTT Gamma Distributed with Invariant model’. De bomen zijn gevisualiseerd met FigTree 1.4.3 en geëxporteerd in pdf-format. In (A) en (B) zijn grote groepen Nederlandse virussen uit het seizoen 2018/2019 geclusterd (‘collapsed’) weergegeven als grote witte driehoeken met vernoeming van het aantal taxa in de tak, waarbij de lengte van de driehoek een maat is voor de diversiteit binnen het cluster. Zwarte driehoekjes en vierkantjes markeren respectievelijk de vaccinstammen van 2018/2019 en 2019/2020. Rode stippen markeren taxa uit het seizoen 2018/2019. In (B) zijn aan de rechterkant diverse clades weergegeven waaraan in de tekst wordt gerefereerd.</span></p>
<p>Moleculaire surveillance van bof-, mazelen- en rodehondvirus</p>
Rogier Bodewes, Rob van Binnendijk, Irene Veldhuijzen
Samenvatting In het huidige Rijksvaccinatieprogramma wordt vaccinatie tegen bof, mazelen en rodehond aangeboden aan alle kinderen op de leeftijd van 14 maanden en 9 jaar. Infecties met voornamelijk bof- en mazelenvirus komen in Nederland (en vele andere landen) echter nog steeds voor. Grote uitbraken van bof vonden plaats tussen 2009 en 2012. De laatste grote uitbraak van mazelen was in 2013-2014 en van rubella in 2004-2005. Jaarlijks zijn er geregeld sporadische gevallen en kleine clusters/uitbraken van mazelen- en bofvirus. Door het systematisch verzamelen van gegevens over het vóórkomen van infecties met deze virussen wordt in kaart gebracht waar en hoe vaak deze ziekten nog voorkomen en kunnen uitbraken gesignaleerd worden. Moleculaire surveillance speelt hier een belangrijke rol in. Hiertoe worden sequenties bepaald van virussen die in verschillende laboratoria zijn gedetecteerd. Door het onderzoeken van (vrijwel) complete genomen van virussen kan de moleculaire resolutie vergroot worden, waardoor op een fijner niveau clustering kan worden aangetoond. Nieuwe sequentietechnieken spelen hier een belangrijke rol in, waarbij de inzet van deze technieken op globale schaal nodig is om de meest bruikbare informatie te verkrijgen.
Abstract According to the current national immunization program in the Netherlands, vaccination against mumps, measles and rubella is offered to all children aged 14 months and 9 years. However, infections with mainly mumps and measles virus still occur in the Netherlands (and many other countries). Major outbreaks of mumps occurred from 2009 to 2012, the last major outbreak of measles was in 2013-2014, and of rubella in 2004-2005. In addition, there are occasional sporadic cases and small clusters/outbreaks of measles and mumps. By systematically collecting data on the occurrence of infections with these viruses, it is mapped where and how often these diseases still occur and outbreaks can be identified. Molecular surveillance plays an important role in this. For this purpose, sequences are determined of viruses detected in different laboratories. For measles and rubella virus, online (closed) databases have been set up by the WHO to which sequences are added worldwide. Molecular resolution can be increased by determining (near) complete genomes of viruses. New sequencing techniques play an important role in this and the use of these techniques on a global scale is crucial to obtain the most useful information.
Introductie Bof, mazelen en rodehond zijn virale aandoeningen, veroorzaakt door drie verschillende virussen. Sinds 1987 is er in Nederland een gecombineerd vaccin beschikbaar waarin levende verzwakte varianten van deze drie virussen zitten. Dit vaccin wordt volgens het huidige schema van het Rijksvaccinatieprogramma aangeboden aan kinderen van 14 maanden en van 9 jaar.
Zowel het bof-, mazelen- als rodehond virus zijn RNA-virussen, maar behoren tot verschillende families. Het bofvirus en mazelenvirus behoren tot de familie van de Paramyxoviridae, terwijl het rodehondvirus tot de familie van de Togaviridae behoort. Genetisch zijn de drie virussen relatief stabiel. Het is dan ook niet nodig dat deze virussen in het vaccin regelmatig aangepast worden aan op dat moment circulerende virusstammen. De levend verzwakte bof-, mazelen- en rodehondvirusvaccinstammen die enkele decennia geleden zijn ontwikkeld, zijn nog steeds in staat om voldoende immuniteit te induceren tegen de virussen die op dit moment circuleren.[1,2]
In het Global Vaccine Action Plan 2011-2020 van de Wereldgezondheidsorganisatie (WHO) is besloten om te streven naar eliminatie van mazelen en rubella in verschillende delen van de wereld. In Nederland komt mazelen buiten de grote uitbraken onder niet-gevaccineerden weinig meer voor, maar wereldwijd overleden er in 2017 nog meer dan 100.000, voornamelijk kinderen.[3] Om de wereldwijde verspreiding van mazelenvirus te stoppen wordt een tweedosisschema aanbevolen, met een vaccinatiegraad van minimaal 95 procent voor beide doses.[1]
Om de impact van het vaccinatieprogramma te evalueren is, naast het bepalen van de vaccinatiegraad, surveillance nodig. De surveillance bestaat uit ziekte- en laboratoriumsurveillance. Door middel van het analyseren van sequentiedata (moleculaire surveillance) van pathogenen kunnen gedetecteerde uitbraken verder gekarakteriseerd en transmissieketens geïdentificeerd worden. Moleculaire surveillance kan laten zien of er in een bepaald land of gebied sprake is van endemische circulatie van een virus of dat er alleen sprake is van importgevallen. In de Nederlandse situatie – met een relatief hoge vaccinatiegraad en lage incidentie - is het belangrijk om van elk sporadisch geval en van minimaal een deel van de gevallen in een nieuwe transmissieketen materiaal te verzamelen voor moleculaire surveillance.
In dit artikel bespreken we de achtergrond van de moleculaire surveillance van bof-, mazelen- en rodehondvirus. Daarnaast geven we een overzicht van de resultaten van de moleculaire surveillance van bof-, mazelen en rodehondvirus in Nederland van de afgelopen jaren. Ook worden op basis van recente data de mogelijkheden en beperkingen van moleculaire surveillance aangegeven.
Surveillance Bof, mazelen en rodehond zijn meldingsplichtige infectieziekten. In het kader van de Wet publieke gezondheid ontvangen GGD’s meldingen van medisch-microbiologische laboratoria en artsen. De GGD voert bron- en contactonderzoek uit en verzamelt aanvullende informatie over onder andere eventuele complicaties, de vaccinatiestatus en de meest waarschijnlijke herkomst van besmetting. Deze gegevens worden met beperkte persoonsgegevens (geslacht, geboortejaar, viercijferige postcode) geregistreerd in Osiris, een onlinesysteem dat beheerd wordt door het RIVM. Met de gegevens uit deze ziektesurveillance wordt de epidemiologie beschreven en wordt onderzocht welke risicofactoren een rol kunnen spelen en welke interventies effectief zijn. Naast de ziektesurveillance wordt materiaal opgevraagd voor typering. Moleculaire surveillance kan informatie geven over de bron en transmissiepatronen. Daarnaast is moleculaire surveillance belangrijk voor de certificering van eliminatie.
De laatste grote uitbraak van mazelen was in 2013-2014, van bof waren er in de periode 2009 tot 2012 achtereenvolgende uitbraken onder studenten (over drie seizoenen), en de laatste grote uitbraak van rubella was in 2004-2005.[4-6] In de periode 2015 tot en met juni 2019 was de incidentie van mazelen minder dan 1 per miljoen inwoners in 2015-2017, 1,4 in 2018 en 5,2 per miljoen in de eerste helft van 2019. Ter vergelijking was de incidentie 160 gevallen per miljoen tijdens de laatste epidemie in 2013. De incidentie van bof varieerde in deze periode tussen 2,7 (2017) en 7,2 per miljoen (eerste helft 2019). Van rubella werd sinds 2015 slechts één geval gemeld.
Endemische transmissie binnen een bepaald gebied is volgens de WHO gedefinieerd als de continue transmissie van een inheems of geïmporteerd mazelen- of rubellavirus gedurende 12 of meer maanden.[7] In Nederland is dat de afgelopen 36 maanden niet aan de orde geweest. Zelfs bij de laatste uitbraak van mazelen in 2013-2014 was het virus binnen 12 maanden geëlimineerd, wat we te danken hebben aan de algeheel hoge graad van immuniteit in de Nederlandse bevolking.
Moleculaire surveillance Moleculaire surveillance van het bof-, mazelen- en rodehondvirus is op dit moment gebaseerd op de sequentie van beperkte delen van de genomen van deze virussen. De te onderzoeken delen van het genoom zijn in eerste instantie geselecteerd op hun relatieve grote genetische variatie ten opzichte van andere delen van het genoom. Aangezien wereldwijd dezelfde delen van het genoom onderzocht worden, kunnen relatief goede vergelijkingen gemaakt worden.
Mazelenvirus Voor de genotypering van het mazelenvirus (15.894 baseparen) beveelt de WHO aan om minimaal de sequentie te karakteriseren van de 450 nucleotiden (N-450) coderend voor de 150 aminozuren van het COOH-einde van het kerneiwit (N). De WHO heeft acht verschillende groepen onderscheiden op basis van genetische verschillen, de groepen A tot en met H. Binnen die groepen worden er op dit moment 24 genotypen onderscheiden, te weten A, B1-3, C1, C2, D1-11, E, F, G1-3 en H1-2.[8] Alle vaccinstammen behoren tot genotype A. Er zijn geen aanwijzingen dat er nog genotype A-wildtype-mazelenvirussen circuleren. In het geval dat genotype A wordt gedetecteerd, bijvoorbeeld tijdens uitbraken, is er sprake geweest van een kort daarvoor uitgevoerde mazelenvaccinatie.
Volgens de richtlijnen van de WHO worden alle door WHO-referentielaboratoria verzamelde sequenties van gedetecteerde mazelenvirussen verstuurd naar de online database Measles Nucleotide Surveillance (MeaNS; www.who-measles.org). Op deze manier ontstaat inzicht in de wereldwijde verspreiding van genetische varianten van mazelenvirussen, al is er sprake van een duidelijke bias aangezien niet in alle landen sequenties worden gekarakteriseerd en/of ingestuurd van gedetecteerde mazelenvirussen.[9]
In tabel 1 wordt een overzicht gegeven van het aantal meldingen van mazelen in Nederland per jaar in de periode 2015 tot en met juni 2019. De patiënten waren tussen de 0 en 57 jaar (mediaan 28). Mazelenvirussen van het genotype B3 en D8 veroorzaken de recente grote uitbraken van mazelen in Europa. Ook de mazelenuitbraak in Nederland in 2013-2014 werd veroorzaakt door het D8-genotype. In de jaren daarna werden in Nederland ook alleen infecties met deze genotypen gerapporteerd. In de periode 2015 tot en met juni 2019 was van 84 van de 98 patiënten een monster beschikbaar voor typering. In 79 gevallen (81 procent) kon ook daadwerkelijk het genotype bepaald worden, 60 daarvan behoorden tot het genotype D8 (figuur 1). Er werden in deze periode 17 epidemiologische clusters gemeld van tussen de twee en elf patiënten.[11] In totaal was 61 procent van de patiënten onderdeel van een cluster, dit aandeel was hoger in de jaren met een groter aantal meldingen (tabel 1). Het merendeel van de patiënten (68 procent) heeft de infectie in het buitenland opgelopen of was gerelateerd aan een importgeval. Van 14 clusters was er van ten minste twee patiënten materiaal voor typering beschikbaar. Vijf clusters waren op basis van een unieke D8-sequentie ook als een apart cluster te onderscheiden, de andere negen clusters waren moleculair niet van elkaar te onderscheiden. De sequenties van 29 van de 36 mazelenvirussen, waarvan in 2019 een genotype gekarakteriseerd kon worden, waren identiek (figuur 2). Dit dendrogram is gemaakt op basis van UPGMA (unweighted pair group method with arithmetic mean) en bevat 60 in Nederland gedetecteerde genotype D8- mazelenvirussen in 2015-2019 (met kleur weergegeven) en 20 epidemiologisch relevante genotype D8- sequentievarianten van mazelenvirus de afgelopen jaren (‘named strains’, wereldwijd; met wit weergegeven). De grootte van de cirkels geeft het aantal virussen weer en de kleuren van de (parten van de) cirkels de jaren waarin die virussen gedetecteerd zijn. Het in grijs weergegeven virus is een genotype D1-mazelenvirus; MVi/Bristol.GBR/0.74, wat gebruikt is als referentiestam voor het bepalen van de phylogenetische afstand tussen de virussen (outgroup). De dendrogram is gemaakt met BioNumerics versie 7.6.3
Uit de data van de moleculaire surveillance van mazelenvirus in Nederland in 2019 valt niet of zeer beperkt op te maken of er sprake is van circulatie van mazelenvirus in Nederland, of van verschillende importgevallen met dezelfde genotypesequentie (figuur 2). De waarde van moleculaire surveillance op basis van een relatief klein deel van het genoom is daarmee te beperkt wanneer uitbraken worden veroorzaakt door één genotypevariant, zoals in 2019 het geval is. Wanneer zo’n genotype bovendien domineert binnen heel Europa kunnen mazelenvirussen die in Nederland worden gedetecteerd bij importgevallen uit verschillende landen toch exact dezelfde sequentie hebben, zonder epidemiologische link. In dit geval kan de resolutie van de moleculaire surveillance vergroot worden door additionele onderdelen van het genoom of zelfs het complete genoom van mazelenvirussen te karakteriseren.[12-14] Op dit moment worden ook op in Nederland gedetecteerde mazelenvirussen verschillende methoden toegepast om sequenties van additionele delen van het genoom te verzamelen om zo de moleculaire resolutie te vergroten. De eerste resultaten geven al aan dat mazelenvirussen van verschillende recente epidemiologische clusters in Nederland door middel van het bepalen van additionele delen van het mazelenvirusgenoom ook moleculair van elkaar te onderscheiden zijn.
Rodehondvirus Net als voor het mazelenvirus is voor rubellavirus (rodehond) een specifiek deel van het genoom geselecteerd voor de genotypering, in dit geval de 739 nucleotiden van het E1-gen (posities 8731-9469). Er zijn op dit moment 13 genotypen bekend, 1a, 1B-1J, 2A-C, waarbij 1a een zogeheten ‘provisional’ genotype is, waartoe het rubellavaccin behoort, maar waarvan wordt aangenomen dat deze als wildtypevirus niet meer circuleert.[15,16] Momenteel circuleren vooral nog genotype 1E en 2B wereldwijd. De laatste kleine uitbraak van rubellavirus van het type 2B was in Nederland in 2013, met meer dan 50 gevallen. In de jaren daarna zijn er in Nederland slechts enkele gevallen van rubellavirus gerapporteerd.[10]
Bofvirus Voor de genotypering van bofvirussen wordt de complete nucleotidensequentie van het zogeheten small hydrophobic (SH) eiwit en de naast gelegen niet-coderende regio’s gebruikt. Dit is het meest variabele deel van het bofvirusgenoom.[17] In totaal zijn dit 316 nucleotiden. Er zijn verschillende primersets die gebruikt kunnen worden voor de genotyperingen van bofvirus. De WHO heeft enige jaren geleden voorgesteld om de gebruikte nomenclatuur voor de genotyperingen van bofvirussen aan te passen. Volgens de nieuwe nomenclatuur worden er nu 12 verschillende genotypen herkend, genaamd A-N met uitzondering van E, M. Daarnaast is er ook voorgesteld om genotype G-bofvirussen verder op te splitsen, gezien de genetische variabiliteit.[17]
Tussen 2009 en 2012 waren er grote uitbraken van bofvirus onder studenten.[5] Deze uitbraken werden vrijwel uitsluitend veroorzaakt door bofvirussen van het genotype G. Ook in de jaren daarna werden bofvirussen van het genotype G het meest gedetecteerd in Nederland. Zo nu en dan zijn er de afgelopen jaren bofvirussen met een ander genotype (C, D, H, K) gedetecteerd, als gevolg van import uit landen waar die genotypen circuleren.[17] Hoewel het SH-gen voor het bofvirus het gebied is met de meeste genetische variatie, is ook deze regio genetisch relatief stabiel. Zo nu en dan zijn er importgevallen van genotype G-bofvirussen uit verschillende genetische clusters, maar het merendeel van de genotype G-bofvirussen die van 2014 tot en met 2019 in Nederland werden gedetecteerd zijn genetisch identiek. Dit suggereert dat in Europa/Nederland al enkele jaren een beperkt aantal bofvirussen circuleert, zie ook het dendrogram in figuur 3. Dit is gemaakt op basis van UPGMA en bevat 111 in Nederland gedetecteerde genotype G-bofvirussen in 2014-2019. De grootte van de cirkels geeft het aantal virussen weer en de kleuren van de (parten van de) cirkels de jaren waarin die virussen gedetecteerd zijn. Het virus weergegeven met de witte cirkel is de referentiestam MuVi/London.GBR/03.02. De dendrogram is gemaakt met BioNumerics versie 7.6.3.
In 2018 en de eerste zes maanden van 2019 waren er negen epidemiologische clusters bestaande uit 2 tot 14 patiënten. Van zes clusters van genotype G-bofvirus was meer dan één monster beschikbaar voor genotypering (totaal 19 monsters). Van bofvirussen gedetecteerd in vier (A-D) van deze clusters waren de SH-sequenties identiek (totaal 13 monsters). Net als bij het mazelenvirus, kan de resolutie van de moleculaire surveillance van bofvirussen vergroot worden door genoominformatie van aanvullende genen en/of regio’s te karakteriseren. Hiervoor kunnen bijvoorbeeld bepaalde niet-coderende gebieden (NCR) tussen het N- en P-gen, het P- en M-gen en het M- en F-gen (N-P, P-M, M-F NCR’s) of de genen coderend voor het HN- en F-eiwit worden gebruikt.[18-20] Uit analyse van de NCR-sequentiedata van de bofvirussen gedetecteerd in de vier clusters met identieke SH-sequentie in 2018-2019, bleek dat er sprake was van enige variatie waardoor virussen uit de verschillende clusters toch genetisch van elkaar onderscheiden kunnen worden (figuur 4). Recent was er in een grote stad sprake van een verhoogd aantal bofgevallen. Hoewel een gemeenschappelijke bron van het bofvirus hier het meest waarschijnlijk leek, bleek op grond van de sequentieanalyse dat er ten minste twee onderscheidende virusvarianten circuleerden. Dit laat zien dat we hier te maken moeten hebben gehad met ten minste twee transmissieketens van het bofvirus.
Toekomst moleculaire typering bof-, mazelen- en rodehondvirus Door de ontwikkeling van nieuwe sequentietechnieken is het eenvoudiger geworden om aanvullende genetische informatie van virale genomen te verzamelen. Hierdoor is het mogelijk de moleculaire resolutie te vergroten en kunnen clusters en individuele gevallen nu beter aan elkaar gelinkt worden of juist worden herkend als afzonderlijke (import)gevallen. Op dit moment zijn er echter toch nog enkele uitdagingen om uit verschillende soorten monsters een betrouwbaar compleet viraal genoom te onderzoeken. Het voorbeeld van bofvirus en de huidige situatie van mazelenvirus in Nederland en Europa geven duidelijk de toegevoegde waarde van het verkrijgen van complete virale genomen aan. Daarnaast is het voor de moleculaire surveillance van mazelen- en bofvirus van groot belang dat in meerdere landen additionele genoominformatie wordt verzameld en dat deze data wordt gedeeld.
Dankwoord Graag willen wij alle medewerkers van laboratoria en GGD’s bedanken voor het insturen van monsters voor genotyperingen. Daarnaast willen we de collega’s van het RIVM bedanken die betrokken zijn geweest bij het karakteriseren van de sequenties van de mazelen- en bofvirussen die gebruikt zijn voor de analyses in dit artikel.
World Health Organization (WHO). Measles vaccines: WHO position paper – April 2017. Weekly Epidemiological Record (WER) 2017;17:205-28.
World Health Organization (WHO). Rubella vaccines: WHO position paper. Weekly Epidemiological Record (WER) 2011;29:301-16.
World Health Organization (WHO). Measles: key facts. 2019. Beschikbaar via https://www.who.int/news-room/fact-sheets/detail/measles. Website bezocht op 18-11-2019.
Woudenberg T, van Binnendijk RS, Sanders EA, et al. Large measles epidemic in the Netherlands, May 2013 to March 2014: changing epidemiology. Euro Surveill. 2017;22(3).
Sane J, Gouma S, Koopmans M, et al. Epidemic of mumps among vaccinated persons, The Netherlands, 2009-2012. Emerg Infect Dis. 2014;20: 643-8.
Hahne S, Macey J, van Binnendijk R, et al. Rubella outbreak in the Netherlands, 2004-2005: high burden of congenital infection and spread to Canada. Pediatr Infect Dis. J 2009;28:795-800.
WHO Regional Office for Europe. Eliminating measles and rubella. Framework for the verification process in the WHO European Region. 2014. Beschikbaar via http://www.euro.who.int/en/health-topics/communicable-diseases/measles-and-rubella/publications/2014/eliminating-measles-and-rubella.-framework-for-the-verification-process-in-the-who-european-region.
Rota PA, Brown K, Mankertz A, et al. Global distribution of measles genotypes and measles molecular epidemiology. J Infect Dis. 2011; 204:S514-23.
The role of extended and whole genome sequencing for tracking transmission of measles and rubella viruses: report from the Global Measles and Rubella Laboratory Network meeting, 2017. Wkly Epidemiol Rec. 2018;93:55-9.
National Institute for Public Health and the Environment (RIVM); Schurink-van 't Klooster TM, de Melker HE (Eds). The National Immunisation Programme in the Netherlands: surveillance and developments in 2017-2018: 2018.
WHO Regional Office for Europe. Surveillance Guidelines for Measles, Rubella and Congenital Rubella Syndrome in the WHO European Region, Updated December 2012, 2012. Beschikbaar via https://apps.who.int/iris/handle/10665/260123.
Penedos AR, Myers R, Hadef B, Aladin F, Brown KE. Assessment of the Utility of Whole Genome Sequencing of Measles Virus in the Characterisation of Outbreaks. PLoS One. 2015;10:e0143081.
Harvala H, Wiman A, Wallensten A, Zakikhany K, Englund H, Brytting M. Role of Sequencing the Measles Virus Hemagglutinin Gene and Hypervariable Region in the Measles Outbreak Investigations in Sweden During 2013-2014. J Infect Dis. 2016;213:592-9.
Sundell N, Dotevall L, Sansone M, et al. Measles outbreak in Gothenburg urban area, Sweden, 2017 to 2018: low viral load in breakthrough infections. Euro Surveill. 2019; 24(17).
Rivailler P, Abernathy E, Icenogle J. Genetic diversity of currently circulating rubella viruses: a need to define more precise viral groups. J Gen Virol. 2017;98:396-404.
Jin L, Orvell C, Myers R, et al. Genomic diversity of mumps virus and global distribution of the 12 genotypes. Rev Med Virol. 2015;25:85-101.
Bodewes R, van Rooijen K, Cremer J, Veldhuijzen IK, van Binnendijk R. Optimizing molecular surveillance of mumps genotype G viruses. Infect Genet Evol. 2019;69:230-4.
Gouma S, Cremer J, Parkkali S, Veldhuijzen I, van Binnendijk RS, Koopmans MPG. Mumps virus F gene and HN gene sequencing as a molecular tool to study mumps virus transmission. Infect Genet Evol. 2016;45:145-50.
Gavilan AM, Fernandez-Garcia A, Rueda A, et al. Genomic non-coding regions reveal hidden patterns of mumps virus circulation in Spain, 2005 to 2015. Euro Surveill. 2018;23(15).
Rijksinstituut voor Volksgezondheid en Milieu (RIVM), Centrum Infectieziektebestrijding - Infectieziekteonderzoek, Diagnostiek en laboratorium Surveillance, Bilthoven, dr. R. Bodewes, wetenschappelijk medewerker. RIVM, Centrum Infectieziektebestrijding – Centrum Immunologie van Infectieziekten en Vaccins, Bilthoven , dr. R. van Binnendijk, wetenschappelijk medewerker. RIVM, Centrum Infectieziektebestrijding – Epidemiologie en Surveillance van Infectieziekten, Bilthoven, dr. I.K. Veldhuijzen, senior epidemioloog. Correspondentieadres: rogier.bodewes@rivm.nl.
<p><span class="Smaller90"><strong>Tabel 1.</strong> Aantal meldingen en kenmerken van mazelenpatiënten per jaar in de periode 2015 t/m juni 2019</span></p>
2015
2016
2017
2018
1e helft 2019
Totaal
Aantal meldingen
7
6
16
24
45
98
Leeftijd (mediaan)
32
20
30
26
24
28
Importstatus*
Import
5
4
6
9
12
36
Importgerelateerd
2
1
3
5
20
31
Onbekend
0
1
7
10
13
31
% import(gerelateerd)
100%
83%
56%
58%
71%
68%
Epidemiologisch cluster
Onderdeel van cluster
0
2
8
17
33
60
Solitair geval
7
4
12
7
12
42
% in cluster
0%
33%
50%
71%
73%
61%
<p><span class="Smaller90"><strong>Figuur 1.</strong> Overzicht van gedetecteerde genotypen bij gerapporteerde mazelengevallen in Nederland van 2015 tot en met juni 2019. Wit: geen genotype bepaald, lichtgrijs: genotype D8, donkergrijs: genotype B3.</span></p>
<p><span class="Smaller90"><strong>Figuur 2</strong>. Dendrogram van recente mazelenvirussen gebaseerd op de nucleotidesequenties van een gedeelte van het N-gen (450 nucleotiden).</span></p>
<p><span class="Smaller90"><strong>Figuur 3</strong>. Dendrogram van recente genotype G-bofvirussen gebaseerd op de nucleotide sequenties van het SH gen (316 nucleotiden). </span></p>
<p><span class="Smaller90"><strong>Figuur 4. </strong>Dendrogrammen van verschillende NCR’s van verschillende bofvirussen van vier epidemiologische clusters (A-D) met identieke SH-sequenties in 2018-2019. Het virus weergegeven met de witte cirkel is de referentiestam MuVi/London.GBR/03.02. De dendrogrammen zijn gemaakt met BioNumerics versie 7.6.3.</span></p>
<p>Visie op toekomstige surveillance van respiratoire bacteriële pathogenen</p>
Rob Mariman, Thijs Bosch, Willem Miellet, Gerlinde Westerhout-Pluister, Janieke van Veldhuizen, Lieke Sanders, Krzysztof Trzciński
Thema: Vaccinaties (2)
2019;27:nr4
Pneumokokken, surveillance, dragerschap
Samenvatting De relatie tussen dragerschap en ziekte kan worden gebruikt voor het monitoren van de effectiviteit van vaccinatieprogramma’s gericht tegen respiratoire bacteriën. De gouden standaard voor dragerschapsdetectie is bacteriekweek uit een nasofaryngeale (pneumokokken) en orofaryngeale (meningokokken) swab. Detectie van deze bacteriën kan worden uitgebreid door het testen van speeksel. De hoge microbiële dichtheid en diversiteit van orale monsters compliceren echter de kweek, waardoor de gevoeligheid van deze methode wordt verminderd. Door gebruik te maken van moleculaire methoden kan de detectie van deze potentiele pathogenen sterk worden verbeterd. Met nieuwe moleculaire diagnostiek hebben we aangetoond dat orofaryngeale swabs superieur zijn in vergelijking met nasofaryngeale swabs voor het aantonen van pneumokokkendragerschap bij volwassenen. Met moleculaire diagnostiek is tevens aangetoond dat speeksel een belangrijk reservoir is voor pneumokokken bij volwassenen, een bevinding die in het begin van de vorige eeuw ook werd waargenomen. Het gebruik van speeksel voor surveillance van pneumokokkenziekte en andere respiratoire bacteriële infecties moet nog verder worden onderzocht.
Aandoeningen aan de luchtwegen, waaronder longontsteking (pneumonie), vormen samen met cardiovasculaire ziekten en kanker de belangrijkste oorzaak van sterfte in Europa. Streptococcus pneumoniae (pneumokok) is de meest voorkomende bacteriële ziekteverwekker van buiten een zorginstelling opgelopen pneumonie (community-acquired pneumonia; CAP). Daarnaast is de pneumokok, ondanks de beschikbaarheid van vaccins, de oorzaak van 75 procent van alle invasieve bacteriële infecties. Wereldwijd overlijden jaarlijks één miljoen volwassenen en 500.000 kinderen (5 procent van de totale kindersterfte) als gevolg van infectie met S. pneumoniae.[1] De hoogste incidentie van invasieve pneumokokkenziekte (IPD) wordt gevonden bij volwassenen boven de 65 jaar, meestal als gevolg van een invasieve pneumonie, vooral bij volwassenen van 75 jaar en ouder.
Ondanks de hoge ziektelast veroorzaakt door pneumokokkenziekte, maken pneumokokken deel uit van de commensale microbiota van de bovenste luchtwegen en vindt transmissie van mens op mens vaak plaatst zonder klinische verschijnselen. Deze vorm van kolonisatie, ook wel dragerschap genoemd, komt het meest voor bij kinderen. Kinderen onder de drie jaar hebben een dragerschapprevalentie van 60 tot 80 procent en zijn hiermee het belangrijkste reservoir van pneumokokken in de populatie.[2] Ondanks de hoge ziektelast bij ouderen werd er tot voor kort nauwelijks dragerschap van de bacterie gerapporteerd in deze groep.
Abstract The link between carriage and disease is utilized in monitoring effects of vaccines targeting respiratory bacteria. The gold standard for carriage detection is the culturing of live bacteria from nasopharyngeal (pneumococcus) or oropharyngeal (meningococci) swabs. These samples can be complemented with collection of saliva. A critical factor for oral samples is the high density and diversity of bacteria which diminishes the sensitivity of culture. Molecular methods have largely improved the detection of pathogens. By applying molecular diagnostics we have reported oropharyngeal swabs in adults to be superior to nasopharyngeal swabs for the detection of pneumococci. More importantly, we detected a large reservoir of pneumococci in the saliva of carriers, a finding in line with century-old studies reporting on high carriage rates. The use of saliva in the surveillance of pneumococcal disease and of other respiratory diseases needs to be explored.
Pneumokokkenvaccins Het belangrijkste antigeen van de pneumokok, een grampositieve diplokok, is het kapsel van polysacharide. Afhankelijk van de samenstelling van verschillende kapselpolysachariden kunnen meer dan 90 serotypen worden onderscheiden. Antistoffen tegen dit kapselantigeen beschermen tegen ziekte. Van alle serotypen is 25 procent verantwoordelijk voor meer dan 80 procent van alle invasieve pneumokokkenziekten. Er zijn momenteel twee typen vaccin beschikbaar, beide op basis van het kapselpolysachariden als antigeen. Het 23-valente polysacharidevaccin bevat kapselantigenen van 23 serotypen. Dit polysacharidevaccin wordt aanbevolen voor ouderen boven de 60 jaar.[3] Het polysacharidevaccin induceert echter een onvoldoende hoge en blijvende immuunrespons bij zuigelingen. Het tweede type vaccin, de polysacharide-eiwitconjugaatvaccins (PCV), induceren wel voldoende beschermende antistoffen en immunologisch geheugen na twee of meer vaccinaties bij hele jonge zuigelingen. Vaccinatie van zuigelingen met PCV’s biedt niet alleen bescherming tegen systemische en lokale infecties veroorzaakt door serotypen die zijn opgenomen in het conjugaatvaccin maar hiermee wordt ook acquisitie van dragerschap met vaccintypen (VT) voorkomen. Sinds de introductie van PCV-7 in 2006 in het Rijksvaccinatieprogramma en de vervanging door PCV-10 in 2011 voor zuigelingen, is in Nederland sprake van een afname van meer dan 90 procent van zowel VT-dragerschap als ziekte.[4] Deze afname wordt niet alleen bij gevaccineerde zuigelingen waargenomen, maar in alle leeftijdsgroepen door preventie van dragerschap en verspreiding van VT’s door kinderen (groepsimmuniteit). De huidige 10-valente en 13-valente PCV’s bieden echter uitsluitende bescherming tegen 10 tot 13 van de meer dan 90 bekende serotypen. Na de introductie van de PCV’s hebben bovendien niet-vaccin serotypen voor een belangrijk deel de ecologische niche ingevuld in dragerschap bij kinderen. Deze niet-vaccin serotypen veroorzaken in toenemende mate ziekte in de populatie, niet zozeer bij kinderen, maar vooral bij ouderen die meer dan 90 procent van alle ziektelast vertegenwoordigen. De zorg is dat deze ‘serotype replacement’ het effect van PCV’s in de bescherming tegen pneumokokkenziekte progressief ondermijnt.
Verbetering van pneumokokkensurveillance Voor een effectief vaccinatiebeleid is anticipatie op veranderingen in serotypen en stammen die verantwoordelijk zijn voor de ziektelast, van cruciaal belang. Beter inzicht in de onderliggende mechanismen van de indirecte gevolgen van vaccinatie, zoals groepsimmuniteit en ‘serotype replacement’, vereisen monitoring van pneumokokken, niet alleen bij patiënten maar ook bij het reservoir van asymptomatische dragers. In Nederland wordt de impact van pneumokokkenvaccinatie geëvalueerd door de monitoring en serotypering van isolaten van 25 procent van alle IPD-patiënten door het Referentie Laboratorium voor Bacteriële Meningitis van het AMC. Daarnaast wordt sinds 2005 pneumokokkendragerschap bij jonge kinderen en hun ouders iedere drie jaar gemonitord met behulp van crosssectionele cohortonderzoeken. Hoewel met deze surveillance van IPD en dragerschap de impact van PCV-vaccinatie goed te volgen is, blijft de huidige surveillance niet toereikend voor een volledig beeld van de dynamiek van pneumokokken onder invloed van vaccinatie en tijd. Het evalueren van bestaande en het opzetten van toekomstige interventiestrategieën voor pneumokokkenziekte vereist accurate data. Dit geldt zowel voor pneumokokkenziekten, waaronder CAP, als voor asymptomatisch dragerschap. Dit moet worden uitgevoerd in alle leeftijdsgroepen, met name ouderen, en in groepen met en zonder verhoogd risico. De gouden standaard van de World Health Organisation (WHO) - het kweken van nasofarynx- (en monsters van orofarynxswab bij volwassenen) - is hiervoor te weinig sensitief, vanwege de lage densiteit van pneumokokkendragerschap bij volwassenen en ouderen.[5] Moleculaire diagnostiek en het gebruik van speeksel als surveillancemiddel geven nieuwe mogelijkheden voor dragerschapsdetectie. Op deze wijze wordt ook bij volwassenen 20 tot 40 procent dragerschap van pneumokokken vastgesteld.[5,6] Om deze reden zet het RIVM in op nieuwe diagnostische methoden met moleculaire technieken, om dragerschap en verspreiding bij volwassenen en vooral ouderen beter te monitoren. Ook is standaardisatie van afname en verwerking van monsters van groot belang voor betrouwbare vergelijking tussen studies en landen.
Maximalisatie van S. pneumoniae-detectie Een van de belangrijkste uitdagingen is het verbeteren van de sensitiviteit en specificiteit van de huidige detectiemethoden voor pneumokokken. Hoewel het kweken en serotyperen van nasofaryngeale uitstrijkjes nog steeds een uitstekende methode is voor het monitoren van dragerschap van pneumokokken bij zuigelingen en jonge kinderen, tonen onze recente studies aan dat de gevoeligheid van deze aanpak snel afneemt met de leeftijd. Dit is vooral het geval bij kinderen van vijf jaar en ouder, omdat de densiteit van dragerschap dan afneemt en de orofarynx een belangrijke niche voor pneumokokkendragerschap wordt.[2] Wij hebben laten zien dat bij volwassenen detectie in speeksel met behulp van moleculaire methoden gevoeliger is dan de ‘gouden standaard’ van de WHO, waarbij wordt gekweekt uit nasofarynx en orofarynx (figuur 1).[5-9] Moleculaire methoden vereenvoudigen daarnaast de determinatie van serotypen en kwantificeren de dynamiek van dragerschap over de tijd. Dit maakt speeksel, dat eenvoudig en niet-invasief kan worden verzameld, zeer geschikt voor dragerschapsurveillance in alle leeftijdsgroepen. Speeksel kan ook worden gebruikt voor immunologische monitoring van IgG- en IgA-antilichamen geïnduceerd na PCV- vaccinatie of na blootstelling door dragerschap. Hoewel er aanwijzingen zijn voor onderdiagnose van S. pneumoniae-geïnduceerde pneumonie vanwege een beperkte detectie van pneumokokken bij volwassenen met de huidige diagnostische testen, moet de eventuele geschiktheid van speeksel voor de surveillance van pneumokokkenziekte en andere respiratoire infecties nog verder worden onderzocht. Studies hiernaar zijn inmiddels opgestart, waarbij samenwerkingen met ziekenhuizen en medisch-microbiologische laboratoria essentieel zijn. Tevens is synchronisatie en integratie van verschillende studies bij patiënten bij wie andere ziekteverwekkers worden bestudeerd, van belang.
Toekomst van surveillance Optimalisatie van interventiestrategieën om luchtweginfecties te reduceren vereisen een integrale aanpak. Daarbij moet de surveillance van S. pneumoniae en andere respiratoire bacteriële pathogenen worden geïntegreerd in kosteneffectiviteitsstudies en impactstudies naar het effect van preventieve en therapeutische interventies. Hierbij dienen andere factoren die bijdragen aan het infectierisico ook meegenomen te worden, zoals leeftijd, comorbiditeit, virale luchtweginfecties, genetica van de gastheer, luchtkwaliteit en sociaaleconomische factoren.
Surveillanceprogramma’s in bestaande cohorten zouden moeten worden uitgebreid met longitudinale dragerschapsstudies, transmissiestudies en studies naar verbeterde diagnostiek bij pneumokokkenpneumonie. Een andere prioriteit is de implementatie van gestandaardiseerde data-analyse voor ‘deep sequencing’ van polymicrobiële monsters en ‘whole genome sequencing‘ (WGS) van individuele bacteriële stammen.
We zijn van mening dat effectieve surveillance in de toekomst berust op het samenbrengen van diverse benaderingen waaronder de huidige conventionele diagnostiek, kweekonafhankelijke moleculaire diagnostiek, WGS voor het in kaart brengen van de populatiestructuur en immunosurveillance van de vaccinrespons. De meest kosteneffectieve aanpak zal afhangen van de specifieke vraagstelling, waarbij rekening dient te worden gehouden met de limitatie van de diverse methoden. Het RIVM heeft meer dan een decennium ervaring in de surveillance van pneumokokkendragerschap onder kinderen en de surveillance van IPD. Onze ambitie is het verder ontwikkelen, valideren en implementeren van innovatieve surveillancemethoden, omdat dit volgens ons de sleutel vormt voor weloverwogen beslissingen over toekomstige vaccinatieprogramma’s.
World Health Organization. International travel and health: pneumococcal disease. http://www.who.int/ith/diseases/pneumococcal/en/
Wyllie AL, Wijmenga-Monsuur AJ, van Houten MA, et al. Molecular surveillance of nasopharyngeal carriage of Streptococcus pneumoniae in children vaccinated with conjugated polysaccharide pneumococcal vaccines. Sci Rep. 2016;6:23809.
Gezondheidsraad. Vaccinatie van ouderen tegen pneumokokken. publicatienr. 2018/05, Den Haag 28 februari 2018.
van Gils EJ, Veenhoven RH, Hak E, et al. Effect of reduced-dose schedules with 7-valent pneumococcal conjugate vaccine on nasopharyngeal pneumococcal carriage in children: a randomized controlled trial. JAMA. 2009;302:159-67.
Trzcinski K, Bogaert D, Wyllie A, et al. Superiority of trans-oral over trans-nasal sampling in detecting Streptococcus pneumoniae colonization in adults. PLoS One. 2013;8:e60520.
Krone CL, Wyllie AL, van Beek J, et al. Carriage of Streptococcus pneumoniae in aged adults with influenza-like-illness. PLoS One. 2015;10(3):e0119875.
Wyllie AL, Rumke LW, Arp K, et al. Molecular surveillance on Streptococcus pneumoniae carriage in non-elderly adults; little evidence for pneumococcal circulation independent from the reservoir in children. Sci Rep. 2016;6:34888.
Bosch A, van Houten MA, Bruin JP, et al. Nasopharyngeal carriage of Streptococcus pneumoniae and other bacteria in the 7th year after implementation of the pneumococcal conjugate vaccine in the Netherlands. Vaccine. 2016;34:531-9.
Rijksinstituut voor Volksgezondheid en het Milieu (RIVM), Centrum voor Infectieziekteonderzoek, Diagnostiek en laboratorium Surveillance, Bilthoven, dr. R. Mariman, dr. T. Bosch, wetenschappelijk medewerkers, W.R. Miellet, promovendus, G.N. Westerhout-Pluister, N.J. van Veldhuizen, analisten, prof. dr. E.A.M. Sanders, chief science officer RIVM. Wilhelmina Kinderziekenhuis/Universitair Medisch Centrum Utrecht, prof. dr. E.A.M. Sanders, kinderarts, hoogleraar Immuundeficiënties en Recidiverende Infecties, dr. K.P. Trzciński, microbioloog, associate professor. Correspondentieadres: K. Trzciński of R. Mariman (K.Trzcinski@umcutrecht.nl of rob.mariman@rivm.nl).
<p><span class="Smaller90"><strong>Figuur 1. </strong>Pneumokokkendragerschap naar leeftijd. Detectie met behulp van de ‘gouden standaard’ - conventionele kweek uit nasofaryngeale (NP) en orofaryngeale (OP) swabs - en moleculaire detectie in speeksel. De streeplijn toont dragerschap bij ouders aan en de doorgetrokken lijn dragerschap bij 20- tot 45-jarigen met minimaal contact met jonge kinderen. [Samengesteld uit referentie 5-9].</span></p>
<p>Vaccinatie tegen meningokokkenziekte</p>
Mirjam Knol, Wieke Freudenburg, Guy Berbers, Helma Ruijs, Hester de Melker, Arie van der Ende
Samenvatting Meningokokkenziekte is een ernstige ziekte die gepaard gaat met hoge mortaliteit en morbiditeit. Serogroep B, C, W en Y veroorzaken nagenoeg alle ziektegevallen in Nederland. Asymptomatisch dragerschap van meningokokken is het hoogst bij adolescenten. Hierdoor spelen zij een belangrijke rol bij de transmissie van meningokokken. Er zijn conjugaatvaccins beschikbaar tegen meningokokken A, C, W en Y, en eiwitvaccins tegen meningokokken B. Vanwege een stijging in meningokokkenziekte C is meningokokken C-vaccinatie ingevoerd in het Rijksvaccinatieprogramma (RVP) in 2002, met tegelijkertijd een inhaalcampagne voor 1- tot 18-jarigen. Dit heeft geleid tot een sterke daling in meningokokkenziekte C bij gevaccineerde en ongevaccineerde leeftijdsgroepen. De incidentie van meningokokkenziekte was het afgelopen decennium historisch laag, maar sinds 2015 neemt de incidentie toe vanwege een stijging in meningokokken W. Vanwege deze toename is het RVP uitgebreid met een quadrivalent meningokokken ACWY-vaccin voor jonge kinderen en adolescenten, en was er in 2018 en 2019 een inhaalvaccinatiecampagne voor adolescenten. Vaccinatie tegen meningokokken B is momenteel geen onderdeel van het RVP vanwege onzekerheid over de effectiviteit van het vaccin in combinatie met het relatief vaak voorkomen van koorts na vaccinatie. Ondanks effectieve vaccinatieprogramma’s komt meningokokkenziekte nog steeds voor en is alertheid bij artsen nodig om deze ernstige ziekte tijdig te herkennen en te behandelen. Daarnaast is een goed surveillancesysteem nodig om uitbraken en trends te detecteren en het RVP te verbeteren.
Abstract Meningococcal disease is a severe disease associated with high mortality and morbidity. Serogroups B, C, W and Y cause nearly all disease in the Netherlands. Asymptomatic meningococcal carriage is highest in adolescents. Therefore, they play an important role in the transmission of meningococci. Conjugate vaccines are available against serogroup A, C, W, and Y, and protein vaccines against serogroup B. Meningococcal C vaccination has been introduced in the Dutch national immunization program (NIP) in 2002, together with a catch-up campaign among 1-18 year olds, after an increase in meningococcal C disease. This led to a strong decrease in meningococcal C disease in vaccinated and unvaccinated age groups. The incidence of meningococcal disease has been historically low in the last 10 years, but since 2015 the incidence has been increasing due to a rise in serogroup W. Therefore, the NIP has been extended with a quadrivalent meningococcal-ACWY-vaccine for young children and adolescents, and a catch-up campaign for adolescents was performed in 2018 and 2019. Meningococcal B vaccination is not included in the Dutch NIP because of uncertainty about vaccine effectiveness and the relatively high prevalence of fever after vaccination. Despite effective vaccination programs, meningococcal disease is still prevalent, and physicians should be aware of the symptoms to recognize and treat meningococcal disease in a timely manner. In addition, a good surveillance system is needed to detect outbreaks and trends, and to improve the NIP.
Inleiding Invasieve meningokokkenziekte is een zeldzame maar zeer ernstige infectieziekte veroorzaakt door Neisseria meningitidis. De afgelopen jaren kwamen er 100 tot 200 ziektegevallen per jaar voor in Nederland, waarvan 5 tot 10 procent overleed. Op basis van het kapsel van de meningokok worden twaalf serogroepen onderscheiden, waarvan B, C, W en Y nagenoeg alle ziektegevallen in Nederland veroorzaken. Sinds 2002 is vaccinatie tegen meningokokken C (MenC) opgenomen in het Rijksvaccinatieprogramma (RVP) op de leeftijd van 14 maanden (tabel 1). Sinds 2018 is deze vaccinatie vervangen door het quadrivalente MenACWY-vaccin, dat bescherming biedt tegen vier serogroepen. Daarnaast zijn in 2018 en 2019 alle 14- tot 18-jarigen opgeroepen voor deze vaccinatie vanwege de toegenomen incidentie van meningokokkenziekte W. Vanaf 2020 wordt MenACWY-vaccinatie opgenomen in het RVP voor kinderen in het jaar waarin ze 14 jaar worden. Er zijn ook vaccins gericht tegen meningokokken B beschikbaar. Deze vaccins zijn in lijn met het advies van de Gezondheidsraad niet opgenomen in het RVP.[1] In dit artikel beschrijven we het ziektebeeld en de epidemiologie van meningokokkenziekte, de verschillende meningokokkenvaccins die beschikbaar zijn en de achtergrond van het vaccinatiebeleid tegen meningokokkenziekte in Nederland.
Meningokokkenziekte: dragerschap, transmissie en ziektebeeld Gezonde mensen kunnen de meningokok bij zich dragen in de neus-keelholte zonder daar ziek van te worden. Dragerschap is relatief laag bij jonge kinderen met een prevalentie van circa 5 procent, neemt toe tijdens de tienerjaren tot een prevalentie van 20 tot 25 procent bij 18- tot 22-jarigen, en daalt daarna weer tot 5 tot 10 procent bij volwassenen.[2,3] Overdracht van de meningokok vindt plaats via direct contact zoals zoenen, maar kan ook plaats vinden via de lucht door aerosolvorming bij hoesten, niezen of praten.[4] Roken, uitgaan, bezoeken van drukke plekken (‘crowding’), intiem contact en het drinken van alcohol zijn risicofactoren voor dragerschap onder adolescenten.[3] Een enkele keer leidt dragerschap tot ziekte waarbij meningitis en/of sepsis kan optreden. Waarom dragerschap bij de ene persoon tot ziekte leidt en bij de andere niet, is niet goed bekend. Meningokokkenziekte kan zeer snel fataal verlopen. Alarmsymptomen van meningokokkenziekte betreffen: een zéér abrupt begin van heftig ziek zijn, koude rillingen, hoge koorts, algehele lichaamspijn, het onvermogen om te kunnen staan, onbegrepen ernstige ongerustheid (bij ouders), diarree, niet wegdrukbare vlekjes/huidbloedinkjes (petechiën). Deze petechiën ontwikkelen zich meestal pas vanaf 8 à 10 uur na eerste klachten en kunnen daarna zeer snel uitbreiden in grootte en aantal. Andere alarmsymptomen zijn tachycardie, vertraagde capillaire refill, acrocyanose, hypotensie, tachypneu, sufheid en – meestal pas na ongeveer 12 tot 24 uur – zich ontwikkelende meningeale prikkeling (luierpijn, nekstijfheid, foto- en acousticofobie). Vroegtijdige herkenning blijft lastig voor huisartsen en spoedeisende hulpartsen, door de veelal aspecifieke symptomen en de zeldzaamheid van de ziekte. Zelfs bij snelle en adequate behandeling overlijdt 5 tot 10 procent van de patiënten. Daarnaast ondervindt circa 6 procent van de patiënten ernstige langetermijncomplicaties zoals gehoorverlies, amputatie van ledematen en hersenschade.[5] Mensen met asplenie, complementdeficiënties, of een open verbinding tussen nasofarynx en meningen hebben een verhoogde kans op meningokokkenziekte. Voor aspleniepatiënten wordt meningokokkenvaccinatie daarom geadviseerd (zie https://lci.rivm.nl/richtlijnen/asplenie). De Nederlandse Vereniging voor Kindergeneeskunde adviseert ook meningokokkenvaccinatie bij afweerstoornissen waaronder complementdeficiënties (zie https://www.nvk.nl/Kwaliteit/Werkboeken/Kinderimmunologie). De meeste patiënten met meningokokkenziekte zijn echter gezonde mensen zonder onderliggend lijden.
Surveillance van meningokokkenziekte Sinds 1905 is meningokokkenziekte een meldingsplichtige ziekte. Melding aan de GGD dient plaats te vinden indien een patiënt ten minste een van de volgende vijf symptomen heeft: koorts, meningeale prikkeling, petechiën, septische shock of septische artritis, in combinatie met laboratoriumbevestiging (positieve kweek of PCR voor N. meningitidis of aantonen van gramnegatieve diplokokken in normaal steriele plaats).[4] Na melding stelt de GGD contactonderzoek in. Aan huisgenoten en zeer nauwe contacten van de patiënt wordt zo spoedig mogelijk antibioticumprofylaxe verstrekt. In de praktijk wordt deze profylaxe soms al door de behandelend arts gegeven bij een (sterke) verdenking op meningokokkenziekte. Wanneer er sprake is van een bewezen geval van meningokokkenziekte door een serogroep A, C, W of Y wordt, naast chemoprofylaxe, vaccinatie van nauwe contacten aangeraden.
Laboratoriumsurveillance wordt uitgevoerd door het Nederlands Referentie Laboratorium voor Bacteriële Meningitis (NRLBM) in het Amsterdam UMC in samenwerking met het RIVM. Het NRLBM ontvangt sinds 1959 op vrijwillige basis isolaten uit bloed of liquor die positief zijn voor N. meningitidis van alle microbiologische laboratoria in Nederland. Van deze isolaten wordt de serogroep bepaald door middel van Ouchterlonygeldiffusie en PCR. Daarnaast vindt typering plaats op basis van DNA-sequentie coderend voor variabele regio’s van de buitenmembraaneiwitten PorA en FetA. Whole genome sequencing wordt toegepast om het sequentietype en het klonale complex te bepalen. Het klonale complex is mogelijk geassocieerd met de mate van transmissie en virulentie.[6] Het NRLBM test ook gevoeligheid voor penicilline en rifampicine.
Epidemiologie van meningokokkenziekte Historische data van de incidentie van meningokokkenziekte laten fluctuaties in de tijd zien: in de zestiger jaren was de incidentie circa 0,5 per 100.000 personen per jaar, terwijl de incidentie in de negentiger jaren 4,0 per 100.000 personen per jaar was.[7] In 2001 nam de incidentie van meningokokkenziekte toe naar 4,5 per 100.000 personen (circa 700 ziektegevallen), veroorzaakt door een toename van serogroep C (figuur 1). Na invoering van MenC-vaccinatie in het RVP in 2002 en de inhaalcampagne onder 1- tot 18-jarigen, daalde de incidentie van meningokokkenziekte C sterk. Sinds 2000 is ook de incidentie van meningokokkenziekte B sterk gedaald van circa 400 ziektegevallen per jaar naar 60 tot 80 per jaar. Waardoor dit komt, is onbekend. Sinds 2015 is er een stijging van meningokokkenziekte-W van minder dan vijf ziektegevallen vóór 2015 naar iets meer dan 100 in 2018. Deze toename wordt veroorzaakt door een klonaal complex 11 (cc11)-stam, die ook in andere Europese landen opkomt.[6,8,9] Meningokokken behorende tot hetzelfde klonale complex veroorzaakten ook de toename van meningokokkenziekte C rond 2000 in Nederland en andere Europese landen. De incidentie van meningokokkenziekte Y is laag met circa 20 ziektegevallen per jaar.
Meningokokkenziekte komt in het algemeen meer voor bij jonge kinderen, met name kinderen jonger dan twee jaar, en bij adolescenten. De leeftijdsverdeling verschilt echter per serogroep (figuur 2). Voor meningokokkenziekte-B is de incidentie verreweg het hoogst bij kinderen tot twee jaar (circa vijf per 100.000 in 2017-2018), gevolgd door kinderen van twee tot vier jaar en adolescenten. Vóór invoering van MenC-vaccinatie had meningokokkenziekte-C eenzelfde leeftijdsverdeling. Meningokokkenziekte-W komt ongeveer evenveel voor bij kinderen tot twee jaar als bij adolescenten. Daarnaast komt meningokokkenziekte-W meer voor bij volwassenen vanaf 45 tot 50 jaar, en met name bij mensen van 80 jaar of ouder. Meningokokkenziekte-Y daarentegen komt weinig voor bij jonge kinderen en adolescenten en juist meer bij volwassenen vanaf 65 jaar.
Ook de mortaliteit varieert per serogroep. Voor serogroep B is de mortaliteit circa 5 procent; in de afgelopen jaren overleden er één tot vijf patiënten per jaar. Sinds de toename van meningokokkenziekte-W in 2015 zijn 45 meningokokken W-patiënten overleden (17 procent); in 2018 overleden 23 patiënten. Met name bij adolescenten is de mortaliteit hoog (25 procent). Voor serogroep Y is de mortaliteit 5 tot 10 procent met nul tot drie gevallen van overlijden per jaar.
Vaccinatie tegen meningokokken C Al sinds 1980 zijn er polysacharidevaccins tegen meningokokken beschikbaar. Deze polysacharidevaccins zijn echter niet effectief bij kinderen, geven maar korte tijd bescherming, en induceren geen immunologisch geheugen en boosterrespons. Sinds 1999 zijn er conjugaatvaccins tegen meningokokken C beschikbaar waarbij het kapselpolysacharide gekoppeld is aan een dragereiwit. Conjugaatvaccins zijn effectiever, werken langer, induceren immunologisch geheugen en zijn al werkzaam vanaf de leeftijd van twee maanden. Conjugaatvaccins beschermen ook tegen dragerschap en reduceren zo de verspreiding van meningokokken.[10,11]
Vanwege de toename van meningokokkenziekte-C in 2001, waarbij er ook meerdere kleine clusters waren, vond er van juni tot december 2002 een grootschalige vaccinatiecampagne plaats.[12] Hierbij werden kinderen van 14 maanden tot en met 18 jaar gevaccineerd met een geconjugeerd MenC-vaccin; tijdens deze campagne werd 94 procent van de kinderen gevaccineerd. Daarnaast werd in september 2002 MenC-vaccinatie op de leeftijd van 14 maanden opgenomen in het RVP. Na invoering van MenC-vaccinatie was er een snelle en sterke afname van het aantal patiënten met meningokokken C te zien, zowel in de gevaccineerde leeftijdsgroepen als in ongevaccineerde leeftijdsgroepen.[10,13] Door vaccinatie van adolescenten, die het vaakst de meningokok bij zich dragen, nam de transmissie van meningokokken C in de gehele bevolking af, wat dus zorgde voor groepsbescherming. Uit serologisch onderzoek onder de Nederlandse bevolking weten we echter dat de huidige adolescenten niet meer voldoende beschermende antistoffen hebben tegen meningokokken C.[14] Beschermende antistoffen zijn namelijk drie tot vijf jaar na de MenC- vaccinatie op 14 maanden verdwenen. Omdat meningokokkenziekte een zeer snel verloop kent, is het noodzakelijk om circulerende antistoffen te hebben en kan niet gewacht worden op de geheugenrespons. Vanwege de lage hoeveelheid beschermende antistoffen bij adolescenten, hebben verschillende Europese landen waaronder het Verenigd Koninkrijk een aantal jaren geleden een booster-MenC-vaccinatie ingevoerd bij adolescenten om de groepsbescherming tegen meningokokken C op peil te houden. In tegenstelling tot jonge kinderen, zijn de beschermende antistoffen bij adolescenten niet verdwenen na vijf jaar; de duur van bescherming is dus veel langer.[15] We verwachten daarom dat de recente ontwikkeling, waarbij MenACWY-vaccinatie van 14-jarigen onderdeel van het RVP wordt (tabel 1), naast bescherming tegen meningokokken W ook bijdraagt aan het behoud van groepsbescherming tegen meningokokken C in Nederland.
Vaccinatie tegen meningokokken W Sinds 2005 zijn er quadrivalente conjugaatvaccins beschikbaar die beschermen tegen meningokokken A, C, W en Y. Deze vaccins zijn geregistreerd op basis van het percentage mensen dat beschermende antistoffen heeft na vaccinatie. Zowel bij jonge kinderen als bij adolescenten geven MenACWY- conjugaatvaccins een goede immuunrespons die ten minste een aantal jaar aanhoudt.[16,17] In een Nederlandse studie onder adolescenten had 95 procent van de deelnemers een jaar na vaccinatie nog beschermende antistoffen tegen de vier serogroepen.[18] Er is nog weinig ervaring met deze vaccins in vaccinatieprogramma’s en dus weinig bekend over de daadwerkelijke effectiviteit tegen meningokokkenziekte. Verwacht wordt dat de effectiviteit vergelijkbaar zal zijn met die van MenC- vaccins, aangezien de werking en immunogeniciteit van deze vaccins vergelijkbaar zijn. In het Verenigd Koninkrijk worden sinds 2015 13- en 14-jarigen gevaccineerd met het MenACWY-vaccin, eveneens vanwege een toename in meningokokkenziekte W. Ook hebben zij in een inhaalcampagne alle 14- tot 18-jarigen en eerstejaarsstudenten opgeroepen voor MenACWY-vaccinatie.[19] De eerste resultaten van de vaccinatiecampagne onder eerstejaarsstudenten zijn positief: een reductie in het aantal meningokokken W-ziektegevallen van 69 procent en geen ziektegevallen bij gevaccineerden bij een vaccinatiegraad van slechts 37 procent.[20]
Vanwege de toename in meningokokkenziekte W in Nederland sinds 2015, worden sinds mei 2018 kinderen van 14 maanden gevaccineerd met een MenACWY-vaccin.[21] Daarnaast vond er in het najaar van 2018 en het voorjaar van 2019 een MenACWY-vaccinatiecampagne plaats voor jongeren van 14 tot 18 jaar (geboortecohorten 2001-2005; tabel 1). Vaccinatie zal directe bescherming bieden aan deze twee leeftijdsgroepen met een relatief hoge incidentie. Daarnaast wordt verwacht dat er groepsbescherming optreedt door het vaccineren van adolescenten, de groep met het hoogste dragerschap en daardoor belangrijk voor transmissie, zodat ook andere leeftijdsgroepen beschermd worden tegen meningokokken W. Deze groepsbescherming werd destijds ook gezien bij de MenC- vaccinatiecampagne. Door in één keer alle adolescenten van 14 tot 18 jaar te vaccineren, is de verwachting dat deze groepsbescherming binnen één tot twee jaar optreedt. Voorlopige cijfers laten zien dat circa 85 procent van de adolescenten de MenACWY-vaccinatie gehaald heeft in 2018 en 2019. In december 2018 adviseerde de Gezondheidsraad om de MenACWY-vaccinatie op de leeftijd van 14 maanden in de toekomst te continueren, in ieder geval zolang de uitbraak van meningokokken W standhoudt.[1] Daarnaast adviseerde de Gezondheidsraad om MenACWY-vaccinatie van adolescenten van 14 jaar op te nemen in het RVP om groepsbescherming tegen meningokokken C en W in de toekomst te behouden. Beide adviezen zijn overgenomen door de minister en worden geïmplementeerd.
Vaccinatie tegen meningokokken B Het is niet mogelijk om een vaccin tegen meningokokken B te ontwikkelen dat zich richt op het kapsel, zoals voor meningokokken A, C, W en Y, omdat het kapselpolysacharide van meningokokken B weinig immunogeen is. Het vertoont namelijk gelijkenis met humane polysacchariden. Daarom zijn tegen meningokokken B andere type vaccins ontwikkeld, gebaseerd op (buitenmembraam) eiwitten die voorkomen op het oppervlak van meningokokken B. Circulerende meningokokken B-stammen brengen deze eiwitten in verschillende mate tot expressie, waardoor deze vaccins niet werkzaam zijn tegen alle meningokokken B-stammen. Tegelijkertijd zijn de eiwitten niet specifiek voor serogroep B, waardoor de vaccins mogelijk ook bescherming kunnen bieden tegen andere serogroepen. Er zijn in Nederland momenteel twee meningokokken B-vaccins beschikbaar. Het 4C-MenB-vaccin (Bexsero) bevat vier antigenen (NHBA, NadA, fHbp en PorA P1.4) en is geregistreerd voor kinderen vanaf twee maanden. Bij kinderen jonger dan twee jaar moeten meerdere primaire doses (twee of drie) worden gegeven en een boosterdosis. Bij kinderen vanaf twee jaar en volwassenen volstaan twee doses. Het MenB-fHbp-vaccin (Trumenba) bevat twee varianten van het fHbp-eiwit en is geregistreerd voor kinderen vanaf 10 jaar.
Voor beide vaccins zijn schattingen gemaakt welk deel van de circulerende MenB-stammen gedekt worden door het vaccin. Uit een onderzoek met data van vijf Europese landen bleek dat het 4C-MenB-vaccin potentieel bescherming biedt tegen 73 tot 87 procent van de MenB-stammen.[22] Voor het MenB-fHbp-vaccin wordt dit percentage geschat op circa 90 procent op basis van ruim 1800 meningokokkenstammen uit de Verenigde Staten en Europa.[23]
Beide vaccins zijn geregistreerd op basis van beschermende antistoffen tegen een aantal referentiestammen. Er is echter nog weinig bekend over de daadwerkelijke effectiviteit en de duur van bescherming. Het 4C-MenB is inmiddels in een aantal Europese landen opgenomen in het vaccinatieprogramma voor baby’s, waardoor er eerste schattingen van de effectiviteit gemaakt zijn. Resultaten uit het Verenigd Koninkrijk, waar het vaccin in 2015 is ingevoerd, laten zien dat het vaccin een bescherming geeft van 65 tot 80 procent.[24,25] In één regio in Quebec, Canada, is in 2014 een vaccinatiecampagne uitgevoerd met het 4C-MenB-vaccin onder alle 0- tot 20-jarigen vanwege een stijgende incidentie van meningokokkenziekte B. Bij mensen van 20 jaar en jonger daalde de incidentie in 2014-2018 met 96 procent ten opzichte van de periode 2006-2014. In andere regio’s in Quebec, waar niet gevaccineerd werd, daalde de incidentie ook, maar niet zo sterk (58 procent), wat duidt op een effect van vaccinatie in de regio met de vaccinatiecampagne. De vaccineffectiviteit werd geschat op 79 procent.
Voor het MenB-fHbp-vaccin zijn nog geen gegevens over effectiviteit beschikbaar. Er wordt verwacht dat MenB-vaccins geen (groot) effect zullen hebben op meningokokkendragerschap en dat daarmee de mate van groepsbescherming van MenB-vaccins beperkt zal zijn.[26,27]
Beide MenB-vaccins zijn veilig bevonden. Wel zijn het reactogene vaccins, wat betekent dat er relatief veel milde bijwerkingen zijn in vergelijking met andere vaccins.[28,29] Het 4C-MenB-vaccin veroorzaakt meer koorts dan andere vaccins, met name als het gelijktijdig wordt gegeven met andere vaccins.[30] Dit kan bij jonge baby’s leiden tot ziekenhuisopname. In het Verenigd Koninkrijk wordt geadviseerd profylactisch paracetamol te geven; dit verlaagt de kans op koorts zonder de immuunrespons te beïnvloeden.[30] Desondanks zijn er na invoering van 4C-MenB-vaccinatie toenames gerapporteerd in het aantal huisartsconsulten en het aantal ziekenhuisopnames als gevolg van koorts bij gevaccineerde kinderen.[31,32]
In december 2018 adviseerde de Gezondheidsraad om vaccinatie tegen meningokokken B niet op te nemen in het RVP.[1] Door de onzekerheid over de mate en duur van bescherming in combinatie met de grotere kans op koorts, kon de Gezondheidsraad geen uitspraak doen over de aanvaardbaarheid van MenB-vaccinatie. Ook de kosteneffectiviteit van MenB-vaccinatie is zeer ongunstig door de huidige lage incidentie, het ontbreken van groepsbescherming en de hoge vaccinprijs. Wanneer er meer gegevens over de effectiviteit beschikbaar zijn, zal MenB-vaccinatie opnieuw beoordeeld worden door de Gezondheidsraad.
In haar artikel bespreekt Bruijning-Verhagen dat op individueel niveau een andere afweging over MenB-vaccinatie gemaakt kan worden.[33] Zij geeft aan dat, ondanks de suboptimale effectiviteit van het vaccin, in het Verenigd Koninkrijk een afname van het aantal MenB-ziektegevallen werd gezien van 50 procent bij zuigelingen één jaar na invoering van MenB-vaccinatie[24] en benoemt dat de kans op koorts na MenB-vaccinatie vergelijkbaar is met die van andere zuigelingenvaccins als MenB-vaccinatie niet gelijktijdig met andere vaccinaties gegeven wordt en profylactisch paracetamol gegeven wordt. Bruijning-Verhagen pleit voor opname van het MenB-vaccin in het RVP, tegen gereduceerde vaccinprijzen maar mét ruimte voor een individuele keuze van ouders.
Het RIVM geeft op de website informatie over MenB-vaccinatie. Mensen kunnen zichzelf of hun kind op eigen kosten laten vaccineren (circa 250,00 euro) via de huisarts of de GGD.[33]
Conclusie Meningokokkenziekte is een zeldzame maar ernstige infectieziekte. Ondanks effectieve vaccinatieprogramma’s komt meningokokkenziekte nog steeds voor en is alertheid van artsen op de symptomen hiervan geboden voor tijdige herkenning (zie de bovengenoemde alarmsignalen). Het verloop van de incidentie van meningokokkenziekte is onvoorspelbaar. Specifieke stammen kunnen opeens opkomen, zoals werd gezien bij meningokokken C in 2001 en bij meningokokken W in 2015, wat ook gepaard kan gaan met verschuivingen in de leeftijdsverdeling of klinische presentatie. Tegelijkertijd kan de incidentie dalen zonder duidelijke oorzaak, zoals bij meningokokken B sinds eind jaren 90. Een goed surveillancesysteem waarin microbiologische, klinische en epidemiologische data gecombineerd worden is van essentieel belang om een toename in ziektegevallen en verschuivingen in karakteristieken tijdig te detecteren. Vroege signalering van een toename in meningokokkenziekte vindt plaats door wekelijkse signalering vanuit het RIVM. Daarnaast is continue monitoring van de ziektelast nodig als input voor vaccinatiebeleid tegen meningokokkenziekte om zo het RVP te optimaliseren.
Gezondheidsraad. Vaccinatie tegen meningokokken. Den Haag; 2018. Report No.: 2018/18.
Christensen H, May M, Bowen L, Hickman M, Trotter CL. Meningococcal carriage by age: a systematic review and meta-analysis. Lancet Infect Dis. 2010;10:853-61.
van Ravenhorst MB, Bijlsma MW, van Houten MA, et al. Meningococcal carriage in Dutch adolescents and young adults; a cross-sectional and longitudinal cohort study. Clin Microbiol Infect. 2017;23:573.e1-573.e7.
LCI Richtlijn - Meningokokken-meningitis en -sepsis (meningokokkenziekte of meningokokkose). Bilthoven: RIVM; Oktober 2013.
Stoof SP, Rodenburg GD, Knol MJ, et al. Disease Burden of Invasive Meningococcal Disease in the Netherlands Between June 1999 and June 2011: A Subjective Role for Serogroup and Clonal Complex. Clin Infect Dis. 2015;61:1281-92.
Krone M, Gray S, Abad R, et al. Increase of invasive meningococcal serogroup W disease in Europe, 2013 to 2017. Euro Surveill. 2019;24(14).
Bijlsma MW, Bekker V, Brouwer MC, Spanjaard L, van de Beek D, van der Ende A. Epidemiology of invasive meningococcal disease in the Netherlands, 1960-2012: an analysis of national surveillance data. Lancet Infect Dis. 2014;14:805-12.
Ladhani SN, Beebeejaun K, Lucidarme J, et al. Increase in endemic Neisseria meningitidis capsular group W sequence type 11 complex associated with severe invasive disease in England and Wales. Clin Infect Dis. 2015;60:578-85.
Knol MJ, Hahne SJM, Lucidarme J, et al. Temporal associations between national outbreaks of meningococcal serogroup W and C disease in the Netherlands and England: an observational cohort study. Lancet Public Health. 2017;2:e473-e482.
Bijlsma MW, Brouwer MC, Spanjaard L, van de Beek D, van der Ende A. A decade of herd protection after introduction of meningococcal serogroup C conjugate vaccination. Clin Infect Dis. 2014;59:1216-21.
Maiden MC, Stuart JM. Carriage of serogroup C meningococci 1 year after meningococcal C conjugate polysaccharide vaccination. Lancet. 2002;359:1829-31.
De Greeff SC, Van Vliet JA, De Melker HE. De huidige vaccinatiecampagne tegen Meningokokken C. Infectieziekten Bulletin. 2002;13:219-23.
de Greeff SC, de Melker HE, Spanjaard L, van den Hof S, Dankert J. [The first effect of the national vaccination campaign against meningococcal-C disease: a rapid and sharp decrease in the number of patients]. Ned Tijdschr Geneeskd. 2003;147:1132-5.
de Voer RM, Mollema L, Schepp RM, et al. Immunity against Neisseria meningitidis serogroup C in the Dutch population before and after introduction of the meningococcal c conjugate vaccine. PLoS One. 2010;5(8):e12144.
van Ravenhorst MB, Marinovic AB, van der Klis FR, et al. Long-term persistence of protective antibodies in Dutch adolescents following a meningococcal serogroup C tetanus booster vaccination. Vaccine. 2016;34:6309-15.
Meningococcal vaccines: WHO position paper, November 2011. Wkly Epidemiol Rec. 2011;86:521-39.
Pellegrino P, Perrone V, Radice S, Capuano A, Clementi E. Immunogenicity of meningococcal quadrivalent (serogroup A, C, W135 and Y) tetanus toxoid conjugate vaccine: systematic review and meta-analysis. Pharmacol Res. 2015;92:31-9.
van Ravenhorst MB, van der Klis FRM, van Rooijen DM, Sanders EAM, Berbers GAM. Adolescent meningococcal serogroup A, W and Y immune responses following immunization with quadrivalent meningococcal A, C, W and Y conjugate vaccine: Optimal age for vaccination. Vaccine. 2017;35:4753-60.
Campbell H, Saliba V, Borrow R, Ramsay M, Ladhani SN. Targeted vaccination of teenagers following continued rapid endemic expansion of a single meningococcal group W clone (sequence type 11 clonal complex), United Kingdom 2015. Euro Surveill. 2015;20(28).
Campbell H, Edelstein M, Andrews N, Borrow R, Ramsay M, Ladhani S. Emergency Meningococcal ACWY Vaccination Program for Teenagers to Control Group W Meningococcal Disease, England, 2015-2016. Emerg Infect Dis. 2017;23(7).
Knol MJ, Ruijs WL, Antonise-Kamp L, de Melker HE, van der Ende A. Implementation of MenACWY vaccination because of ongoing increase in serogroup W invasive meningococcal disease, the Netherlands, 2018. Euro Surveill. 2018;23(16).
Vogel U, Taha MK, Vazquez JA, et al. Predicted strain coverage of a meningococcal multicomponent vaccine (4CMenB) in Europe: a qualitative and quantitative assessment. Lancet Infect Dis. 2013;13:416-25.
McNeil LK, Donald RGK, Gribenko A, et al. Predicting the Susceptibility of Meningococcal Serogroup B Isolates to Bactericidal Antibodies Elicited by Bivalent rLP2086, a Novel Prophylactic Vaccine. MBio. 2018;9(2).
Parikh SR, Andrews NJ, Beebeejaun K, et al. Effectiveness and impact of a reduced infant schedule of 4CMenB vaccine against group B meningococcal disease in England: a national observational cohort study. Lancet. 2016;388:2775-82.
Minutes of the JCVI meeting in October 2018.
Read RC, Dull P, Bai X, et al. A phase III observer-blind randomized, controlled study to evaluate the immune response and the correlation with nasopharyngeal carriage after immunization of university students with a quadrivalent meningococcal ACWY glycoconjugate or serogroup B meningococcal vaccine. Vaccine. 2017;35:427-34.
Soeters HM, Whaley M, Alexander-Scott N, et al. Meningococcal Carriage Evaluation in Response to a Serogroup B Meningococcal Disease Outbreak and Mass Vaccination Campaign at a College-Rhode Island, 2015-2016. Clin Infect Dis. 2017;64:1115-22.
Flacco ME, Manzoli L, Rosso A, et al. Immunogenicity and safety of the multicomponent meningococcal B vaccine (4CMenB) in children and adolescents: a systematic review and meta-analysis. Lancet Infect Dis. 2018;18:461-72.
Shirley M, Taha MK. MenB-FHbp Meningococcal Group B Vaccine (Trumenba((R))): A Review in Active Immunization in Individuals Aged >/= 10 Years. Drugs. 2018;78(2):257-268.
Prymula R, Esposito S, Zuccotti GV, et al. A phase 2 randomized controlled trial of a multicomponent meningococcal serogroup B vaccine (I). Hum Vaccin Immunother. 2014;10:1993-2004.
Harcourt S, Morbey RA, Bates C, et al. Estimating primary care attendance rates for fever in infants after meningococcal B vaccination in England using national syndromic surveillance data. Vaccine. 2018;36:565-71.
Murdoch H, Wallace L, Bishop J, Robertson C, Claire Cameron J. Risk of hospitalisation with fever following MenB vaccination: self-controlled case series analysis. Arch Dis Child. 2017;102:894-8.
Bruijning-Verhagen P. Advies Gezondheidsraad over meningokokkenvaccinatie. Ned Tijdschr Geneeskd 2019;163.
Rijksinstituut voor Volksgezondheid en Milieu, Centrum voor Infectieziektenbestrijding (CIb), Bilthoven, dr. M.J. Knol, epidemioloog, dr. G.A.M. Berbers, biochemicus, dr. W.L.M. Ruijs, arts infectieziekte bestrijding, dr. H.E. de Melker, epidemioloog. Amsterdam UMC, Nederlands Referentie Laboratorium voor Bacteriële Meningitis (NRLBM), Amsterdam, dr. W. Freudenburg, arts-microbioloog, dr. A. van der Ende, moleculair bioloog. Correspondentieadres: Mirjam Knol (mirjam.knol@rivm.nl)
<p><span class="Smaller90"><strong>Figuur 1.</strong> Incidentie van meningokokkenziekte naar serogroep, 1992 tot 2019* (*t/m juni).</span></p>
<p><span class="Smaller90"><strong>Figuur 2.</strong> Incidentie van meningokokkenziekte naar serogroep en leeftijd, 2017-2018.</span></p>
<p><span class="Smaller90"><strong>Tabel 1.</strong> Vaccinatie tegen meningokokken in Nederland</span></p>
Jaar
Vaccin
Leeftijd
Eenmalig
RVP
2002
MenC-conjugaat
14 maanden-18 jaar
X
2002
MenC-conjugaat
14 maanden
X
2018
MenACWY-conjugaat
14 maanden
X
Okt-nov 2018
MenACWY-conjugaat
13-14 jaar
(cohort mei-dec 2004)
X
Mrt-jun 2019
MenACWY-conjugaat
13-18 jaar
(cohort 2001-2005)
X
2020
MenACWY-conjugaat
14 jaar
X
<p>Ruim 25 jaar Hib-vaccinatie in Nederland</p>
Mirjam Knol, Hester de Melker, Helma Ruijs, Guy Berbers, Wieke Freudenburg, Lieke Sanders, Arie van der Ende
Samenvatting Invasieve Haemophilus influenzae type b (Hib) kan zich manifesteren als meningitis, epiglottitis en pneumonie, wat kan leiden tot levenslange restverschijnselen of overlijden. Hib-vaccinatie wordt sinds 1993 gegeven binnen het Rijksvaccinatieprogramma en is zeer effectief gebleken. De incidentie van invasieve Hib-ziekte is met meer dan 90 procent gedaald na invoering van vaccinatie. Toch zijn er elk jaar nog enkele tientallen, meest ongevaccineerde, ziektegevallen, en zijn er af en toe lichte toenames in de incidentie. Met de invoering van maternale kinkhoestvaccinatie eind 2019 zal het Hib-vaccinatieschema veranderen van een 2-3-4-11-maandenschema naar een 3-5-11-maandenschema indien de moeder tijdens de zwangerschap tijdig gevaccineerd is. Ook andere typen Hi kunnen invasieve ziekte veroorzaken. Van die andere typen komt niet-typeerbare Hi (ntHi) momenteel het meeste voor, met name bij prematuren, immuungecompromitteerden en ouderen. De incidentie van invasieve ntHi-ziekte is in de afgelopen 10 jaar verdubbeld zonder duidelijk aanwijsbare oorzaak. Deze recente ontwikkelingen laten zien dat continue monitoring van invasieve Hi-ziekte belangrijk blijft, om veranderingen in incidentie tijdig te identificeren en de effecten van het vaccinatieprogramma te evalueren.
Abstract Invasive Haemophilus influenzae type b (Hib) disease can present as meningitis, epiglottitis and pneumonia, which can lead to lifelong sequelae or death. Hib-vaccination has been part of the Dutch National Immunization Program since 1993 and has shown to be very effective. The incidence of invasive Hib disease has decreased with more than 90 per cent after introduction of vaccination. Still, every year there are a few dozen Hib cases, mostly unvaccinated, and there have been periods with slight increases in incidence. With introduction of maternal pertussis vaccination at the end of 2019, the Hib-vaccination schedule will change from a 2-3-4-11 month schedule to a 3-5-11 month schedule for children whose mother has been vaccinated on time during pregnancy. Also other types of Hi can cause invasive disease. Of these other types, non-typable Hi (ntHi) currently causes most disease, especially among premature children, immunocompromised patients and elderly. The incidence of invasive ntHi disease has doubled in the last 10 years, without knowing what causes this increase. These recent developments show the importance of continuous monitoring of invasive Hi disease to timely identify changing epidemiology, and to evaluate the effects of the vaccination program.
Inleiding De gramnegatieve bacterie Haemophilus influenzae (Hi) kan ernstige invasieve infecties veroorzaken zoals meningitis, epiglottitis en sepsis, maar veel vaker veroorzaakt Hi (niet-invasieve) luchtweginfecties zoals otitis media, sinusitis en pneumonie. Op basis van het kapselpolysacharide worden zes serotypen onderscheiden: a tot en met f. Ook ongekapselde Hi, zogenoemde niet-typeerbare Hi (ntHi) kunnen (invasieve) ziekte veroorzaken. Sinds de jaren 90 is een vaccin tegen één Hi-type, type b (Hib), beschikbaar. Destijds was Hib een van de belangrijkste veroorzakers van bacteriële meningitis. In Nederland is Hib-vaccinatie in 1993 ingevoerd in het Rijksvaccinatieprogramma voor zuigelingen, ter preventie van ernstige Hib-infecties als meningitis en epiglottitis. Sindsdien is het aantal mensen met invasieve Hib-ziekte sterk afgenomen, van 500 tot 800 per jaar naar circa 40 per jaar, waarvan ongeveer 10 gevaccineerde kinderen per jaar.[1] Daarnaast zijn er nog ruim 150 patiënten met invasieve Hi veroorzaakt door een ander type dan Hib.
In dit artikel beschrijven we het ziektebeeld en de epidemiologie van invasieve Hib-ziekte. Ook het huidige vaccinatiebeleid en recente ontwikkelingen op het gebied van Hib worden besproken. Naast Hib besteden we ook aandacht aan andere Hi-typen.
Invasieve Hib-ziekte: transmissie, ziektebeeld en diagnostiek Hib is een commensaal van de bovenste luchtwegen en wordt verspreid door niezen, hoesten en nauw contact. De bacterie kan ernstige invasieve ziekte veroorzaken. De belangrijkste klinische manifestaties van invasieve Hib-infecties zijn meningitis, epiglottitis, pneumonie en septische artritis.[2] Symptomen van meningitis bij kinderen zijn hoofdpijn, nekstijfheid, braken, koorts en sufheid, terwijl bij zuigelingen het klinisch beeld minder specifiek is. Epiglottitis, een acute ontsteking met zwelling van het strottenhoofdklepje, wordt gekenmerkt door een acuut begin van een infectie met hoge koorts, keelpijn en ernstige slikklachten. Er ontstaat een progressieve benauwdheid bij inademen. Epiglottitis is levensgevaarlijk door acuut verstikkingsgevaar. Hib is tevens een belangrijke verwekker van pneumonie waarbij vaak pleuravocht aanwezig is en symptomen als tachypneu, hoesten, benauwdheid en koorts. Ook kan Hib septische artritis veroorzaken. Bewegingsbeperking, pijn en zwelling van een gewricht en koorts zijn belangrijke symptomen. Andere invasieve ziektebeelden die door Hib veroorzaakt kunnen worden, zijn osteomyelitis, pericarditis, cellulitis en sepsis of bacteriëmie zonder focus. De mortaliteit van invasieve Hib-ziekte is 2 tot 7 procent en restverschijnselen als gehoorverlies, hydrocephalus en epilepsie treden op bij 8 tot 15 procent van de patiënten.[1,3] Mensen met asplenie hebben een hoger risico op Hib-ziekte en een ernstiger verloop van Hib-ziekte. Bij deze patiënten wordt (extra) Hib-vaccinatie geadviseerd (zie LCI-richtlijn Asplenie).
Op basis van het klinische beeld kan meningitis of pneumonie door Hib niet worden onderscheiden van andere bacteriële verwekkers. Een Hi-infectie kan worden vastgesteld door middel van kweek of PCR in materiaal afkomstig uit een normaal steriele plaats. Het bepalen van het Hi-type wordt gedaan door het Nederlands Referentie Laboratorium voor Bacteriële Meningitis (NRLBM) in het Amsterdam UMC.
Hib-vaccinatie Het Hib-vaccin is een conjugaatvaccin waarbij een kapselpolysacharide is gekoppeld aan tetanus toxoid als dragereiwit. Dit vaccin geeft een T-celafhankelijke immuunrespons waardoor er al op jonge leeftijd beschermende antistoffen worden geïnduceerd. Ook wordt er immunologisch geheugen opgewekt waardoor er een boosterrespons optreedt na het herhalen van de vaccinatie. Daarnaast verlaagt Hib-vaccinatie de acquisitie van Hib waardoor Hib-dragerschap, en vervolgens transmissie van Hib, sterk vermindert na vaccinatie.[4] Er bestaan losse Hib-vaccins en combinatievaccins. In het huidige Rijksvaccinatieprogramma wordt Hib-vaccinatie gegeven op de leeftijd van 2, 3, 4 en 11 maanden in een combinatievaccin DTaP-IPV-Hib-HepB, wat ook bescherming biedt tegen difterie, tetanus, kinkhoest, polio en hepatitis B. Sinds invoering van Hib-vaccinatie in 1993 zijn er verschillende wijzigingen geweest in het vaccinatieschema en het gebruikte vaccin (tabel 1). Na invoering van maternale kinkhoest vaccinatie eind 2019 zal het vaccinatieschema opnieuw veranderen (zie Huidige ontwikkelingen). De Hib-vaccinatiegraad ligt sinds de invoering van het vaccin rond de 95 procent.[5]
Surveillance van Hib Laboratoriumsurveillance voor Hi wordt uitgevoerd door het NRLBM in het Amsterdam UMC in samenwerking met het RIVM. Het NRLBM ontvangt (op vrijwillige basis ingestuurde) isolaten van normaal steriele materialen die positief zijn voor Hi, van alle microbiologische laboratoria in Nederland. Van deze isolaten wordt het serotype bepaald door middel van co-agglutinatie. Het serotype kan ook bepaald worden door middel van PCR in niet-gekweekte materialen.
Sinds 2009 is Hib een meldingsplichtige ziekte. Melding aan de GGD dient plaats te vinden indien er sprake is van een ziektebeeld passend bij Hib (meningitis, epiglottitis, bacteriëmie zonder focus of sepsis, artritis, pneumonie, osteomyelitis, pericarditis, cellulitis) en het aantonen van Hib in materiaal afkomstig uit een normaal steriele plaats.[2] Na melding stelt de GGD contactonderzoek in om te beoordelen of het toedienen van profylaxe aan contacten nodig is. Profylaxe met rifampicine is aanbevolen voor alle huisgenoten indien er in het huishouden kinderen jonger dan vier jaar zijn die niet of nog onvolledig gevaccineerd zijn of indien er een immuungecompromitteerd kind deel uitmaakt van het huishouden. In de melding worden verder klinische gegevens zoals ziektebeeld, overlijden en onderliggend lijden, en vaccinatiegegevens gerapporteerd door de GGD.
In een recente analyse door het RIVM bleek dat slechts de helft van de patiënten met invasieve Hib-ziekte die bij het NRLBM bekend zijn, door de GGD gemeld worden.[6] Melden van Hib aan de GGD is van belang voor adequate advisering over chemoprofylaxe ter preventie van ernstige ziekte bij contacten (zie LCI richtlijn) en versterkt de monitoring van Hib door volledigheid van het Nederlandse Hib-surveillancesysteem.
Epidemiologie Hib-ziekte In de periode voor invoering van Hib-vaccinatie in 1993 was Hib één van de belangrijkste veroorzakers van bacteriële meningitis bij kinderen.[7] Bij het NRLBM kwamen in 1992 isolaten van bijna 300 patiënten met invasieve Hib-ziekte binnen; 95 procent van deze patiënten was jonger dan vijf jaar. Dit aantal is waarschijnlijk een onderschatting van het werkelijke aantal Hib-patiënten in Nederland om twee redenen: 1) de dekking van het NRLBM was naar schatting 80 procent voor Hib-meningitis,[8] en 2) in die tijd werden met name isolaten uit liquor ingestuurd en dus geeft dit een onderschatting van het aantal Hib-patiënten met een ander ziektebeeld dan meningitis. Een artikel van Bol uit 1991[1] geeft daarom een schatting van 800 invasieve Hib-patiënten per jaar voor invoering van Hib-vaccinatie.
Na invoering van Hib-vaccinatie daalde de incidentie van invasieve Hib-ziekte echter snel en eind jaren 90 werden er minder dan 20 patiënten per jaar gerapporteerd door het NRLBM, een daling van meer dan 90 procent. In de afgelopen 20 jaar fluctueerde het aantal Hib-patiënten tussen 20 en 50 per jaar (incidentie: 0.2 per 100,000 per jaar) met af en toe een hogere incidentie zoals in 2004/2005 en in 2016-2018 (figuur 1).
In het post-vaccinatietijdperk is nog maar een derde van de Hib-patiënten jonger dan vijf jaar, maar de incidentie is toch nog steeds het hoogst in deze leeftijdsgroep (1,5 per 100.000 per jaar) (figuur 2). Daarna is de incidentie het hoogst onder mensen van 65 jaar of ouder met circa 10 ziektegevallen per jaar (0,3 per 100.000 per jaar). Het overlijdenspercentage ligt ongeveer op 5 procent. Van de kinderen onder de vijf jaar met Hib is circa 60 procent gevaccineerd.[5] Dit is beduidend lager dan de vaccinatiegraad in de populatie (circa 95 procent). Hieruit kunnen we afleiden dat het vaccin een effectiviteit heeft van rond de 90 procent. Een vergelijking van 41 gevaccineerde en 10 ongevaccineerde kinderen met Hib die tussen 2005 en 2013 gediagnosticeerd waren, liet geen significante verschillen zien in onderliggend lijden, overlijden, of restverschijnselen tussen gevaccineerde en ongevaccineerde kinderen.[3]
Andere Hi-serotypen De incidentie van invasieve ziekte door andere gekapselde Hi-bacteriën is laag (figuur 1). Het meest voorkomende type is Hi-type f met 10 tot 20 ziektegevallen per jaar, van wie ongeveer de helft 65 jaar of ouder is. Hi-type a, d en e samen veroorzaken minder dan 10 ziektegevallen per jaar.
De incidentie van invasieve ziekte door niet-typeerbare Hi (ntHI) is de laatste decennia echter toegenomen met momenteel circa 170 ziektegevallen (incidentie: 1,0 per 100.000 per jaar) (figuur 1). In tegenstelling tot invasieve Hib-ziekte, die veelal gezonde kinderen treft, komt invasieve ntHi-ziekte voornamelijk voor bij kwetsbare groepen zoals prematuur geboren kinderen, immuungecompromitteerde patiënten en ouderen.[9] De incidentie is het hoogst bij mensen van 65 jaar en ouder, gevolgd door kinderen jonger dan vijf jaar (figuur 3). Een recente studie vond een mortaliteit van 12 procent, met de hoogste mortaliteit bij patiënten van 80 jaar of ouder (31 procent) en een mortaliteit van 4 procent bij kinderen jonger dan vijf jaar.[9] De klinische manifestatie verschilde per risicogroep; bij neonaten was bacteriëmie zonder focus het meest voorkomende ziektebeeld, bij kinderen jonger dan vijf jaar was dit met name meningitis en bij ouderen met name pneumonie.
Huidige ontwikkelingen
Nederland ten opzichte van andere Europese landen Ondanks de sterke daling van het aantal Hib-ziektegevallen na invoering van vaccinatie en onze hoge vaccinatiegraad, is de incidentie van Hib in Nederland twee tot tien keer hoger dan in andere Europese landen.[10] De afgelopen vijf jaar was de incidentie in Nederland rond de 0,2 per 100.000 ten opzichte van 0,02 per 100.000 in de EU (op basis van 19 rapporterende landen), en bijvoorbeeld 0,03 per 100.000 in het Verenigd Koninkrijk, en 0,1 per 100.000 in Noorwegen en Denemarken. Ook het aantal gevaccineerde kinderen met Hib-ziekte in Nederland (circa 10 per jaar) is hoger dan bijvoorbeeld in Engeland en Wales (zes gevaccineerde Hib-patiënten in de periode 2012 tot 2016).[11] Deze verschillen lijken niet verklaard te kunnen worden door het vaccinatieschema, vaccin of surveillancesysteem; het is vooralsnog onduidelijk wat de oorzaak van deze verschillen dan wel is.
Stijgende incidentie tussen 2011 en 2016 Van 2011 tot 2016 was er een toename in het aantal Hib-ziektegevallen, met name bij kinderen jonger dan vijf jaar. Opvallend was dat deze toename samenviel met de invoering van het DTaP-IPV-Hib-HepB-vaccin voor alle kinderen in Nederland (tabel 1). In een patiënt-controleonderzoek is daarom de effectiviteit van het Hib-vaccin berekend bij kinderen gediagnosticeerd tussen 2003 en 2016.[12] Hieruit bleek dat de vaccineffectiviteit tussen 2011 en 2016 niet lager was dan daarvoor, en dat de vaccineffectiviteit van zowel het DTaP-IPV-Hib-HepB-vaccin als het DTaP-IPV-Hib-vaccin hoog was, respectievelijk 94 en 92 procent. Wel lijkt de effectiviteit af te nemen met de leeftijd, wat een proxy is voor tijd-sinds-vaccinatie, van 95 procent bij kinderen van nul tot twee jaar naar 70 procent bij kinderen van drie en vier jaar; dit was voor beide vaccins het geval.
Vooralsnog is er geen verklaring gevonden voor de tijdelijke stijging van de incidentie in de periode van 2011 tot 2016. In 2017 en 2018 lijkt deze stijging gestopt te zijn. Ook in 2004/2005 was er een periode van onverklaarde stijging van de incidentie.
Verandering in vaccinatieschema Vanaf eind 2019 krijgen alle zwangere vrouwen in Nederland vaccinatie tegen kinkhoest (door middel van DTaP of DTaP-IPV-vaccin) aangeboden op het consultatiebureau. Door de bescherming door maternale antistoffen bij de pasgeborenen kan bij a-term geboren kinderen de eerste vaccinatie tegen kinkhoest uitgesteld worden tot drie maanden en kunnen er twee in plaats van drie primaire doses gegeven worden.[13] Dit geldt alleen indien de moeder tijdens de zwangerschap tijdig (minimaal twee weken voor de bevalling) gevaccineerd is. In het nieuwe schema krijgen kinderen het DTaP-IPV-Hib-HepB-combinatievaccin op de leeftijd van 3, 5 en 11 maanden aangeboden (tabel 1). Premature kinderen, kinderen van wie de moeder positief is voor hepatitis B, en kinderen van wie de moeder niet (tijdig) gevaccineerd is tegen kinkhoest krijgen een extra vaccinatie als ze twee maanden oud zijn. De verwachting is dat de verandering van het schema niet leidt tot meer Hib-ziektegevallen omdat dit schema in veel andere landen al met succes wordt toegepast. Toch blijft continue monitoring van de Hib-incidentie en de effectiviteit van Hib-vaccinatie van belang om eventuele veranderingen tijdig te detecteren.
Andere Hi-typen Een toename van ntHi wordt ook in andere Europese landen en in de Verenigde Staten gezien, ook vooral onder ouderen.[14,15] Vergrijzing van de populatie en een toenemend gebruik van immunosuppressiva dragen mogelijk bij aan de toename. Dat deze veroorzaakt wordt door typevervanging vanwege invoering van Hib-vaccinatie lijkt niet plausibel, omdat de ntHi-patiënten andere klinische karakteristieken hebben dan Hib-patiënten voor invoering van vaccinatie (meer premature kinderen, immuungecompromitteerden en ouderen).
Net als in Nederland lijkt ziekte door andere gekapselde Hi-typen ook in andere Europese landen niet toe te nemen.[15] De Verenigde Staten rapporteren wel een toename van ziekte door Hi-type a.[14] Er wordt gewerkt aan vaccins tegen ntHi en Hi-type a, maar deze zijn nog in (een vroege fase van) ontwikkeling.[16,17] Er is recent een fase 2-klinische studie gedaan naar een ntHi-eiwitvaccin bij volwassenen met COPD.[18] Dit betreft een vaccin bestaande uit drie antigenen: vrij Protein D (PD), wat als conjugaat gebruikt wordt in het 10-valente pneumokokkenconjugaatvaccin, en gefuseerd protein E (PE) en pilin A (pilA). Dit vaccin liet een goede immunogeniciteit zien en een acceptabel veiligheids- en reactogeniciteit-profiel. Vervolgonderzoek moet laten zien of het vaccin daadwerkelijk ziekte door ntHI kan voorkomen.
Conclusie Invasieve Hib-ziekte is een ernstig ziektebeeld waaraan patiënten kunnen overlijden of levenslang restverschijnselen kunnen overhouden. Sinds de invoering van Hib-vaccinatie in 1993 is de incidentie van invasieve Hib-ziekte laag, maar wel hoger dan in andere Europese landen. De effectiviteit van Hib-vaccinatie is hoog. Toch zijn er jaarlijks een aantal gevaccineerde en ongevaccineerde kinderen met Hib-ziekte. Ziekte door niet-typeerbare Hi neemt toe en is met name een probleem bij kwetsbare groepen en ouderen. Surveillance van Hi ziekte is en blijft belangrijk om het vaccinatieprogramma te evalueren en trends te detecteren om zo het Rijksvaccinatieprogramma verder te optimaliseren.
Bol P. [Epidemiology of Haemophilus influenzae type b infections in The Netherlands and elsewhere]. Ned Tijdschr Geneeskd. 1991;135:7-9.
Landelijk Coördinatiecentrum Infectieziektenbestrijding. Haemophilus influenzae type b-infectie. Available from: https://lci.rivm.nl/richtlijnen/haemophilus-influenzae-type-b-infectie. Accessed at
Monge S, Mollema L, de Melker H, Sanders E, van der Ende A, Knol M. Clinical Characterization of Invasive Disease Caused by Haemophilus influenzae Serotype b in a High Vaccination Coverage Setting. J Pediatric Infect Dis Soc. 2018.
Barbour ML. Conjugate vaccines and the carriage of Haemophilus influenzae type b. Emerg Infect Dis. 1996;2:176-82.
The National Immunisation Programme in the Netherlands. Surveillance and developments in 2017-2018. 2018. Report No.: 2018-0124. Available from: https://www.rivm.nl/bibliotheek/rapporten/2018-0124.pdf.
Van den Boogaard J, Van der Ende A, Westerhof A, Ruijs WLM, De Melker HE, Knol MJ. Voldoen medisch microbiologische laboratoria en de GGD aan de landelijke meldingsplicht voor invasieve Hib-ziekte? Infectieziektenbulletin. 2019;30(3).
Spanjaard L, Bol P, de Jong MC, Zanen HC. Bacterial meningitis in 366 children in the Netherlands, 1982-1983. Epidemiology and antibiotic therapy. Tijdschr Kindergeneeskd. 1986;54:1-8.
Spanjaard L, Bol P, Ekker W, Zanen HC. [Incidence of bacterial meningitis in the Netherlands; comparison of 3 registration systems, 1977-1982]. Ned Tijdschr Geneeskd. 1985;129:355-9.
van Wessel K, Rodenburg GD, Veenhoven RH, Spanjaard L, van der Ende A, Sanders EA. Nontypeable Haemophilus influenzae invasive disease in The Netherlands: a retrospective surveillance study 2001-2008. Clin Infect Dis. 2011;53(1):e1-7.
Control ECfDPa. Surveillance Atlas of Infectious Diseases. Available from: https://atlas.ecdc.europa.eu/public/index.aspx. Accessed at 2019.
Collins S, Litt D, Almond R, et al. Haemophilus influenzae type b (Hib) seroprevalence and current epidemiology in England and Wales. J Infect. 2018;76:335-41.
Monge S, Hahne SJ, de Melker HE, Sanders EA, van der Ende A, Knol MJ. Effectiveness of the DTPa-HBV-IPV/Hib-vaccine against invasive Haemophilus influenzae type b disease in the Netherlands (2003-16): a case-control study. Lancet Infect Dis. 2018;18:749-57.
Gezondheidsraad. Vaccinatieschema zuigelingen na maternale kinkhoestvaccinatie. Den Haag; 2018. Report No.: 2018/27.
Soeters HM, Blain A, Pondo T, et al. Current Epidemiology and Trends in Invasive Haemophilus influenzae Disease-United States, 2009-2015. Clin Infect Dis. 2018;67:881-9.
Whittaker R, Economopoulou A, Dias JG, Bancroft E, Ramliden M, Celentano LP. Epidemiology of Invasive Haemophilus influenzae Disease, Europe, 2007-2014. Emerg Infect Dis. 2017;23:396-404.
Murphy TF. Vaccines for Nontypeable Haemophilus influenzae: the Future Is Now. Clin Vaccine Immunol. 2015;22:459-66.
Barreto L, Cox AD, Ulanova M, Bruce MG, Tsang R. The emerging Haemophilus influenzae serotype a infection and a potential vaccine: Implementation science in action. Can Commun Dis Rep. 2017;43:85-8.
Wilkinson TMA, Schembri S, Brightling C, et al. Non-typeable Haemophilus influenzae protein vaccine in adults with COPD: A phase 2 clinical trial. Vaccine. 2019;37:6102-11.
Rijksinstituut voor Volksgezondheid en Milieu (RIVM), Centrum Infectieziektenbestrijding (CIb), Bilthoven. M.J. Knol, epidemioloog, dr. H.E. de Melker, epidemioloog, dr. W.L.M. Ruijs, arts infectieziektenbestrijding, dr. G.A.M. Berbers, biochemicus, prof. dr. E.A. Sanders, kinderarts-infectioloog. Nederlands Referentielaboratorium voor Bacteriële Meningitis (NRLBM), Amsterdam UMC, Amsterdam, dr. W. Freudenburg, arts-microbioloog, dr. A. van der Ende, moleculair bioloog. Correspondentieadres: M.J. Knol (Mirjam.knol@rivm.nl).
<p><span class="Smaller90"><strong>Tabel 1.</strong> Vaccinatieschema en vaccin sinds invoering van Hib-vaccinatie in 1993; vetgedrukte tekst geeft de veranderingen aan.</span></p>
Jaar
Vaccinatieschema
Vaccin
1993
3, 4, 5 en 11 maanden
Los Hib-vaccin
1999
2, 3, 4 en 11 maanden
Los Hib-vaccin
2002
2, 3, 4 en 11 maanden
Combinatievaccin DTwP-IPV-Hib
2004
2, 3, 4 en 11 maanden
Combinatievaccin DTaP-IPV-Hib
2006
2, 3, 4 en 11 maanden
Combinatievaccin DTaP-IPV-Hib-HepB voor kinderen met verhoogd risico op hepatitis B
2011
2, 3, 4 en 11 maanden
Combinatievaccin DTaP-IPV-Hib-HepB voor alle kinderen
Eind 2019
3, 5 en 11 maanden voor kinderen van wie de moeder tijdig gevaccineerd is tegen kinkhoest* tijdens de zwangerschap
2, 3, 5 en 11 maanden voor kinderen van wie de moeder niet gevaccineerd is tegen kinkhoest tijdens de zwangerschap en voor premature kinderen
Combinatievaccin DTaP-IPV-Hib-HepB
<p><span class="Smaller90"><strong>Figuur 1.</strong> Aantal ziektegevallen van invasieve <em>H. influenzae</em>-ziekte naar serotype tussen 1992 en 2018.</span></p>
<p><span class="Smaller90"><strong>Figuur 2.</strong> Aantal ziektegevallen van invasieve <em>H. influenzae</em> type b-ziekte naar leeftijd tussen 2009 en 2018.</span></p>
<p><span class="Smaller90"><strong>Figuur 3. </strong>Aantal ziektegevallen van invasieve niet-typeerbare H. influenzae ziekte naar leeftijd tussen 2009 en 2018.</span></p>
<p>HPV-vaccinatie in Nederland<br /> <span class="Smaller90">10 jaar in het Rijksvaccinatieprogramma, een overzicht</span></p>
Joske Hoes, Tessa Schurink-van ’t Klooster, Hella Pasmans, Pascal van der Weele, Kahren van Eer, Petra Woestenberg, Birgit van Benthem, Hans Boogaards, Fiona van der Klis, Audrey King, Hester de Melker
Thema: Vaccinaties (2)
2019;27:nr4
HPV, vaccinaties, Rijksvaccinatieprogramma
Samenvatting Een persistente infectie met humaan papillomavirus (HPV) kan op termijn leiden tot verschillende soorten (pre-)maligne laesies in de anogenitale en orofaryngeale streek. Sinds 2006 zijn er drie verschillende profylactische vaccins geregistreerd die bescherming bieden tegen hoogrisicotypen HPV. Deze vaccins hebben de potentie om de door HPV veroorzaakte ziektelast sterk te verminderen. In Nederland is sinds 2010 het bivalente HPV-vaccin opgenomen in het Rijksvaccinatieprogramma voor 12-jarige meisjes. De vaccins zijn veilig en zeer effectief in de bescherming tegen (persisterende) hoogrisico-HPV-infecties, zoals ook blijkt uit verschillende Nederlandse onderzoeken. Wanneer in 2023 de eerste gevaccineerde cohorten het bevolkingsonderzoek baarmoederhalskanker instromen, worden naar verwachting ook de eerste effecten op ziekteniveau ((pre)kanker laesies) zichtbaar. In juni 2019 bracht de Gezondheidsraad een advies uit om, naast meisjes, ook jongens te vaccineren tegen HPV en om het vaccin voortaan aan te bieden rond de leeftijd van negen jaar. Tevens adviseert de Gezondheidsraad om volwassenen tot en met 26 jaar de kans te geven zich alsnog te laten vaccineren. In september 2019 besloot het ministerie van Volksgezondheid, Welzijn en Sport het advies van de Gezondheidsraad over te nemen met ingang van 2021.
Abstract A persisting infection with Human Papillomavirus (HPV) can eventually lead to premalignant lesions on the anogenital and oropharyngeal sites. From 2006 onwards, three prophylactic vaccines have been registered for the protection against high-risk types HPV. These vaccines have the ability to strongly reduce the disease burden caused by HPV. In the Netherlands, the bivalent HPV-vaccine was introduced into the National Immunization Program in 2010 as a girls-only vaccine for 12-year-olds. The HPV-vaccines have been shown to be safe and highly effective against (persistent) high-risk HPV infections, as is also evidenced by multiple Dutch studies. When the first vaccinated birth cohorts enter the cervical cancer screening program in 2023, the first effects on disease outcomes are expected to be visible. In June 2019, the Dutch Health council advised to start vaccinating boys in addition to girls and to change the age of vaccination to 9 years. Moreover, the Health Council advices to offer adults up to 26 years of age the opportunity to get the HPV-vaccine through a supplemental vaccination program. In September 2019, the Ministry decided to follow the advice from the Health Council starting from 2021.
Inleiding HPV-infectie is de veroorzaker van baarmoederhalskanker en van andere HPV-gerelateerde kankers aan de penis, anus, vulva en orofarynx. Sinds 2006 zijn vaccins beschikbaar die bescherming geven tegen verschillende hoogrisicotypen HPV. Sinds 2010 is het bivalente HPV16/18-vaccin opgenomen in het Rijksvaccinatieprogramma (RVP) voor 12-jarige meisjes. In dit artikel wordt achtereenvolgens ingegaan op het humaan papillomavirus, de pathogenese van het virus, de vaccins, opname van het bivalente vaccin en monitoring hiervan in het RVP en onderzoek.
HPV Een infectie met het humaan papillomavirus (HPV) is, met een geschat percentage van 80 procent van de mensen die ooit een infectie doormaken, de meest voorkomende seksueel overdraagbare infectie.[1] HPV is een klein, dubbelstrengs DNA-virus uit de Papillomaviridae-familie en kent veel verschillende typen, waarvan er tot nu toe meer dan 200 zijn gekarakteriseerd.[2] De typen worden gegroepeerd binnen verschillende genera, waarvan het alfagenus bestaat uit ongeveer 40 typen die de genitale slijmvliezen kunnen infecteren.[3] Er wordt onderscheid gemaakt tussen hoog- en laagrisicotypen HPV op basis van oncogeen potentieel; van 14 typen (HPV16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66 en 68) is bekend dat zij kanker kunnen veroorzaken, waarvan baarmoederhalskanker de meest voorkomende anogenitale kanker is.[3,4] Daarnaast betreft het ook een deel van de penis-, anus-, vagina- en vulvakankers en van de kankers in het hoofd-halsgebied. Naast hoogrisicotypen zijn er ook laagrisicotypen HPV binnen het alfa-genus die geassocieerd zijn met het ontwikkelen van genitale wratten en laaggradige laesies. HPV6/11 zijn de typen die hierin de grootste bijdrage hebben, met een geschatte attributie van 90 procent aan alle genitale wratten.[5]
Pathogenese De pathogenese voor het verloop van HPV-infectie tot aan maligniteit is het beste beschreven voor baarmoederhalskanker. Een aanhoudende, transformerende infectie met een hoogrisicotype HPV (hrHPV) is een noodzakelijke voorwaarde voor het ontwikkelen van baarmoederhalskanker. HPV kan de basale lagen van het plaveiselepitheel van de cervix infecteren, waarvoor microlaesies (die veelal ontstaan gedurende geslachtsgemeenschap) in het epitheel nodig zijn. Een productieve infectie ontstaat wanneer virusdeeltjes worden uitgescheiden naar het epitheel oppervlak.[6] Ongeveer 80 tot 90 procent van de productieve infecties wordt geklaard door het immuunsysteem, waarbij het virus niet meer gedetecteerd wordt in achtereenvolgende metingen, en verloopt symptoomloos.[7] Virale latentie en mogelijk ook reactivatie bij veranderende omstandigheden kan hierbij niet worden uitgesloten.[5]
In sommige gevallen kan een HPV-infectie aanhouden (persisteren) en veranderen in een transformerende infectie, waarbij voorloperstadia van baarmoederhalskanker kunnen ontstaan, die CIN-laesies (cervicale intra-epitheliale neoplasie) worden genoemd. De early genes van het virus spelen hierbij een belangrijke rol, waarbij E2 vooral betrokken is in de fase van replicatie en de regulatie van E6 en E7, die gezien worden als de virale oncogenen die de celproliferatie stimuleren en herstel voorkomen. De deregulering van deze genen kan zorgen voor een opeenstapeling van genetische fouten, die kunnen leiden tot virale integratie in het genoom van de gastheer, iets wat vaak wordt gezien in baarmoederhalskanker.[5,7]
Er worden laaggradige (CIN1 en deel CIN2) en hooggradige (deel CIN2 en CIN3) afwijkingen onderscheiden, waarbij de hooggradige laesies worden beschouwd als voorloperstadia van cervixcarcinoom.[8] Regressie kan te allen tijde optreden door klaring van het virus, maar hoe hoger de graad van de laesie, hoe kleiner deze kans. De tijd die nodig is om een hooggradige laesie te ontwikkelen wordt geschat op enkele jaren, terwijl voor het ontwikkelen van maligniteit vanuit de primaire infectie meer tijd nodig is; zo’n 12 tot 15 jaar.[9] Ongeveer 70 procent van alle baarmoederhalskanker worden veroorzaakt door HPV16/18.[10] In Nederland worden jaarlijks tussen de 700 en 800 vrouwen gediagnosticeerd met baarmoederhalskanker en overlijden ongeveer 200 vrouwen aan deze vorm van kanker. Tevens kunnen andere anogenitale kankers, waaronder peniskanker (circa 160 nieuwe gevallen per jaar), anuskanker (circa 200 tot 300), vaginakanker (circa 60) en vulvakanker (circa 300 tot 400) en mond- en keelholtekankers (circa 1600 tot 1800) worden veroorzaakt door een aanhoudende hrHPV-infectie (figuur 1), waarbij HPV16 ook bij deze vormen van kanker het meest voorkomt. In tegenstelling tot baarmoederhalskanker is bij de andere genoemde kankervormen een HPV-infectie niet altijd de aanleiding en spelen andere factoren een rol in de pathogenese.[11,12]
Epidemiologie HPV komt veel voor en besmetting vindt plaats door direct epitheelcontact, waarbij seks de belangrijkste manier van overdracht is. HPV is zeer besmettelijk en mede daardoor is penetratie geen noodzakelijke voorwaarde voor infectie; contact van de genitale zones kan voldoende zijn. Een toename in de prevalentie van HPV wordt gezien vanaf het begin van de seksuele activiteit. Voor vrouwen met normale cytologie ligt de piekprevalentie tussen de 20 en 25 jaar, waarna deze met toenemende leeftijd afneemt.[3] Voor mannen lijkt de prevalentie stabieler gedurende het leven, hoewel daarover minder bekend is. Tevens zijn de internationale verschillen in prevalentie groot, met hogere schattingen voor Afrika, Oost-Europa en Zuid-Amerika vergeleken met Westerse landen en Azië.[13] Risicofactoren voor het oplopen van een infectie zijn vooral gerelateerd aan seksueel gedrag, zoals het aantal sekspartners gedurende het leven.[14]
HPV-vaccinatie Sinds 2006 zijn er verschillende profylactische vaccins voor primaire preventie tegen HPV geregistreerd, die gebaseerd zijn op het L1-eiwit (manteleiwit) van het virus. Op basis van virus-like particles (VLP’s) in het vaccin (aangevuld met adjuvans), wordt een immunologische respons en geheugen geïnduceerd zonder dat er sprake is van infectie.[15] Door te vaccineren vóór seksueel debuut wordt een optimale bescherming bewerkstelligd. De vaccins verschillen in aantal HPV-typen waartegen zij bescherming bieden; het bivalente vaccin (Cervarix, GSK) is gericht tegen HPV16/18. In het quadrivalente vaccin (Gardasil, MSD) zijn daar ook de twee meest voorkomende laagrisicotypen HPV6 en 11 aan toegevoegd. Beide vaccins bieden daarnaast kruisbescherming tegen enkele andere hrHPV-typen. Enkele jaren later is een nonavalent vaccin op de markt gebracht (Gardasil9, MSD, 2015), dat niet alleen bescherming biedt tegen HPV6/11/16/18 maar ook tegen hoogrisicotypen HPV31/33/45/52/58. HPV-vaccinatie werd oorspronkelijk geregistreerd voor bescherming tegen baarmoederhalskanker, maar inmiddels is dit uitgebreid naar anus-, vagina- en vulvalaesies. De vaccins kunnen gebruikt worden voor kinderen van negen jaar en ouder. Initieel werd een driedosesschema geadviseerd, maar na een advies van de European Medicines Agency (EMA) in 2014 werd dit aangepast naar een tweedosesschema. Uit immunobridgingsstudies bleek dat de immunogeniciteit onder jonge meisjes (9 tot 14 jaar) die met twee doses waren gevaccineerd, niet inferieur was aan de immunogeniciteit in een oudere populatie (15 tot 25 jaar) die gevaccineerd was met drie doses en bij wie een zeer hoge effectiviteit waargenomen werd.[16,17] Het principe van immunobridging gaat ervan uit dat bij een niet-inferieure immunologische respons, de effectiviteit ook niet inferieur zal zijn. Bij de start van het vaccinatieschema op 15-jarige leeftijd of later wordt geadviseerd alsnog een driedosesschema aan te houden (0, 1, 6 maanden).
Over het therapeutische effect van de huidige vaccins bij een bestaande infectie is tot op heden weinig bekend; de vaccins werken het beste wanneer ze gegeven worden vóór blootstelling aan het virus en zijn ook met dit doel ontwikkeld. Binnen de RCT’s zijn wel deelnemersgroepen geïncludeerd die niet HPV-negatief waren bij aanvang (de zogeheten ‘intention to treat populations’). De vaccineffectiviteit in deze groepen ligt over het algemeen lager.[18,19] Daarnaast wordt in Nederland momenteel de VacCIN-studie uitgevoerd (Aduvant VACcination against HPV in surgical treatment of CIN-lesions, a Randomized Controlled Trial), waarin wordt onderzocht of HPV-vaccinatie na een chirurgische behandeling (LEEP; Loop Electrosurgical Excision Procedure) recidiverende voorloperstadia van baarmoederhalskanker vermindert.
HPV-vaccinatie binnen het RVP In 2008 heeft de Gezondheidsraad advies uitgebracht en geadviseerd HPV-vaccinatie ter preventie van baarmoederhalskanker op te nemen in het Rijksvaccinatieprogramma.[20] Na een inhaalcampagne in 2009 voor meisjes uit de geboortecohorten 1993 t/m 1996, worden meisjes sinds 2010 uitgenodigd voor vaccinatie in het jaar dat zij 13 worden. Zowel bij de inhaalcampagne als in het reguliere vaccinatieprogramma is tot nu toe gebruikgemaakt van het bivalente HPV-vaccin. In eerste instantie kregen meisjes drie doses aangeboden, maar naar aanleiding van het advies van de EMA is dit in 2014 aangepast naar twee doses. Meisjes uit geboortecohort 2001 waren hiermee de eersten die in aanmerking kwamen voor vaccinatie met twee doses (0, 6 maanden).
Monitoring van de HPV-vaccinatiegraad en veiligheid Sinds de invoering van de HPV-vaccinatie in het RVP in 2010 is de uptake een kritische factor gebleken. Hoewel de vaccinatiegraad de eerste jaren stijgende was (van 56 procent in 2010 naar 61 procent in 2013/14), heeft zich de afgelopen jaren een vrij abrupte daling voorgedaan. De vaccinatiegraad kwam daarmee voor 2016 (geboortecohort 2003) en 2017 (geboortecohort 2004) uit op 45,5 procent (figuur 2). Voorlopige cijfers laten een lichte stijging zien voor 2018, die mogelijk gerelateerd is aan de grootschalige meningokokkenvaccinatiecampagne onder jongeren en/of aan de toenemende aandacht voor vaccinatiebereidheid in de media in het algemeen.
Onderzoek onder ouders van meisjes die in aanmerking kwamen voor vaccinatie maar zich niet lieten vaccineren, toonde aan dat zij zorgen hadden over de veiligheid van dit vaccin.[21] Ook in andere landen, waaronder Denemarken en Ierland, werd een sterke daling gezien van de vaccinatiegraad. Deze daling werd vooral ingegeven door veiligheidsoverwegingen van ouders en door media-aandacht daarvoor. Na onder andere een actiever social mediabeleid vanuit het Ierse Nationaal Immunisatie Bureau, het oprichten van een HPV-vaccinatie-alliantie en een uitgebreide mediacampagne nam de bereidheid om te vaccineren in Ierland weer toe.[22]
In Nederland worden de veiligheid en de mogelijke bijwerkingen van HPV-vaccinatie continu gemonitord. Hierover is recent een overzichtsartikel verschenen van Schurink-van ‘t Klooster et al. [23] Het veiligheidsprofiel in Nederland is in lijn met dat wat in buitenlands onderzoek wordt geobserveerd. Er treden milde, voorbijgaande klachten op na de vaccinatie waarbij pijn op de injectieplaats, spierpijn en verminderd gebruik van de arm het meest frequent worden gerapporteerd. In overeenstemming met buitenlandse studies is geen associatie tussen HPV-vaccinatie en eerder gesuggereerde bijwerkingen gevonden, zoals langdurige vermoeidheidsklachten.[24]
Vaccineffectiviteit De lange periode tussen infectie en de ontwikkeling van (pre)kankerlaesies maakt dat de vaccineffectiviteit (VE) tegen kanker pas vele jaren na de start van het vaccinatieprogramma kan worden aangetoond. In de tussentijd vormt de VE tegen aanhoudende infecties een goede graadmeter (intermediate endpoint). Daarom is met de implementatie van de HPV-vaccinatie een Nederlandse prospectieve cohortstudie gestart gericht op het monitoren van de HPV-incidentie in gevaccineerde en ongevaccineerde groepen. Meisjes die in aanmerking kwamen voor HPV-vaccinatie in de inhaalcampagne (geboortecohorten 1993-1994) werden in 2009 en 2010 uitgenodigd voor deelname aan de HAVANA-studie: HPV Among Vaccinated And Non-vaccinated Adolescents. Eenmaal vóór eventuele vaccinatie en ieder jaar daarna leverden de deelneemsters een cervixborstel in waaruit de HPV-infectiestatus op typeniveau werd bepaald en vulden zij een vragenlijst in.[25] Uit deze studie blijkt dat de VE zeer hoog is; er werd een VE van 95 procent tegen persisterende infecties met vaccintypen HPV16/18 gezien tot in ieder geval zeven jaar na vaccinatie (figuur 3). Ook werd significante kruisbescherming gevonden tegen persisterende HPV31/33/45 infecties (66 procent).[11,26]
Tevens is op basis van de HAVANA-data aangetoond dat kortdurende en persistente infecties met HPV16/18 die optreden bij gevaccineerde personen een significant lagere virale load hebben in vergelijking met HPV16/18-infecties bij niet-gevaccineerde personen. Voor infecties met HPV31/45, waar kruisbescherming tegen is gevonden, werd geen lagere virale load gevonden bij gevaccineerde ten opzichte van ongevaccineerde personen. Hoewel de mechanismen van de adaptieve immuniteit op dit moment nog niet duidelijk zijn, suggereert dit dat vaccinatie de kans op langdurende infectie met HPV16/18 zou kunnen verkleinen en ook dat het effect van vaccinatie verder gaat in het beheersen van de infectie na acquisitie.[27]
Ook in ander Nederlands, crosssectioneel onderzoek toonde men de hoge effectiviteit van het bivalente vaccin aan, onder 16- tot 24-jarige bezoekers van centra seksuele gezondheid (voormalig soa-poli’s).[28] In de PASSYON-studie (PApillomavirus Surveillance among STI clinic YOungsters in the Netherlands) daalde de HPV16/18-prevalentie onder alle deelnemende vrouwen van 23 procent voor introductie van vaccinatie in 2009 naar 15 procent in 2015. Ook onder heteroseksuele mannen was een daling zichtbaar (van 17 procent naar 11 procent), wat wijst op groepsbescherming.[29] Daarnaast kon binnen dit onderzoek een VE tegen anale HPV-infecties bij vrouwen worden berekend; hieruit bleek dat de VE tegen anale HPV16/18-infecties 90 procent is, wat een vergelijkbare effectiviteit van het vaccin suggereert als tegen genitale infecties.[30] Tevens is aan de hand van PASSYON-data gekeken naar de relatie tussen type-specifieke VE tegen niet-vaccin-hrHPV-typen en fylogenetische afstand tot HPV16/18. Hieruit bleek dat kruisbescherming tegen hrHPV beter verklaard kon worden aan de hand van genomische afstand tot HPV16/18 (zoals bepaald met whole genome sequencing) dan aan de hand van de afstand gebaseerd op L1-manteleiwitten. Hiermee kan worden gekeken naar de verschillende hoogrisicotypen HPV waarvoor wel of geen kruisbescherming kan worden verwacht op basis van de fylogenie.[31]
Ook in een recente systematische review en meta-analyse met 60 miljoen personen kwam het positieve effect van de introductie van HPV-vaccinatieprogramma’s duidelijk naar voren; acht jaar na invoering van het bivalente en quadrivalente vaccin kwamen HPV16/18-infecties 83 procent en HPV31/33/45-infecties 54 procent minder voor onder meisjes van 13 tot 19 jaar. Daarnaast was de prevalentie van CIN2-laesies sterk gedaald en was er sprake van een vermindering van genitale wratten bij zowel mannen als vrouwen.[32]
Serologie Uit verschillende monitoringsstudies is gebleken dat de immunogeniciteit van het bivalente vaccin zeer hoog is. Zowel gegevens uit de HAVANA-studie (deelneemsters gevaccineerd met drie doses in de catch-upcampagne) als uit de zogeheten HPV2D-studie (deelneemsters gevaccineerd met twee doses binnen reguliere programma) laten hoge antistofresponsen zien tegen vaccintypen HPV16/18 tot respectievelijk negen en vier jaar na vaccinatie. Ook na één dosis is het vaccin immunogeen, maar zowel de antistofconcentraties als de B- en T-celresponsen zijn lager.[33]
Karakterisering van HPV16/18 Recente karaktersering van de DNA-sequentie van het hele genoom van HPV16/18-virussen die bij Nederlandse vrouwen voorkomen, heeft geleid tot nieuwe inzichten in genoomdiversiteit van deze virussen. Er blijkt een grote populatie van HPV16/18-varianten virussen natuurlijk voor te komen bij vrouwen. Op het niveau van het gehele genoom blijkt binnen deze studies bijna iedere vrouw een unieke HPV16- of HPV18-variant te hebben.[34,35] Daarnaast is bij vrouwen met een langdurige HPV16/18-infectie de hele genoomvariantsequentie tot drie jaar na de infectie volledig identiek. Hoe deze enorme virusdiversiteit ontstaat is nog niet duidelijk. Deze bevinding maakt het echter mogelijk om onderscheid te maken tussen primaire en terugkerende HPV16/18-infecties die bijvoorbeeld CIN2/3-laesies veroorzaken.[36]
Langetermijnprojectie op ziektelast De langetermijneffecten van vaccinatie zijn geschat aan de hand van wiskundige modellen waarmee zowel de verspreiding van het virus als de ontwikkeling van kanker werden gesimuleerd. Twee onafhankelijke studies voorspelden een halvering van de incidentie van baarmoederhalskanker bij een min of meer constante vaccinatiegraad van 50 tot 60 procent onder meisjes.[37,38] Bij deze vaccinatiegraad zou de HPV-gerelateerde ziektelast bij mannen met ongeveer een derde afnemen. Interessant is dat het indirecte effect van meisjesvaccinatie volgens modelberekeningen sterker uitvalt bij heteroseksuele mannen dan bij ongevaccineerde vrouwen.[39] Dit is te verklaren doordat indirecte bescherming van ongevaccineerde vrouwen een afspiegeling is van de indirecte bescherming van ongevaccineerde mannen (eerste-orde-effect), en derhalve een tweede-orde-effect is. Deze modelvoorspelling strookt met recente bevindingen in de PASSYON-studie.[29] Hierin is wel een daling van HPV16/18-prevalentie onder heteroseksuele mannen te zien, maar (nog) niet onder ongevaccineerde vrouwen.
Efficiëntie van seksespecifieke vaccinatie Een andere bevinding van modelstudies is dat het verhogen van de vaccinatiegraad onder meisjes een verhoudingsgewijs sterker effect heeft op het voorkomen van HPV-infectie, en daarmee op preventie van HPV-gerelateerde ziekte, dan vaccinatie van jongens.[39] Niettemin is deze asymmetrie mild: voor elk extra meisje zouden twee jongens moeten worden gevaccineerd om een gelijke gezondheidswinst te behalen.[40] Daarnaast wordt de relatieve efficiëntie van beide strategieën ook bepaald door de moeite die het kost om extra meisjes dan wel jongens te includeren.[41] Een gecombineerde strategie biedt meer mogelijkheden op het elimineren van vaccintypen: bij meisjesvaccinatie zou de vaccinatiegraad meer dan 90 tot 95 procent moeten zijn om eliminatie van HPV16 te bereiken, maar bij sekseneutrale vaccinatie is dit volgens modellen mogelijk bij 80 procent onder zowel jongens als meisjes.[42]
Vaccinatie en screening In 2023 stromen de eerste gevaccineerde cohorten uit de catch-upcampagne het bevolkingsonderzoek baarmoederhalskanker (BVO) in. Dit biedt de mogelijkheid om het effect van vaccinatie te meten in (pre)kankerlaesies, aangezien er dan informatie beschikbaar zal zijn over zowel vaccinatiestatus als screeningsresultaat. Dat de gunstige resultaten van bescherming tegen persisterende HPV-infectie zullen resulteren in een goede bescherming tegen voorloperstadia van baarmoederhalskanker, toont onder meer recent onderzoek in Schotland aan. Hierin werden de eerste screeningsuitslagen van (bivalent) gevaccineerde en ongevaccineerde 20-jarige vrouwen vergeleken en bleek dat onder gevaccineerden het voorkomen van CIN3, CIN2 en CIN1 met respectievelijk 89 procent, 88 procent en 79 procent was afgenomen.[43] Toch tonen dergelijke resultaten naast de positieve effecten van vaccinatie, ook de noodzaak van een continue evaluatie van het screeningsprogramma aan. Dit dient optimaal ingericht te zijn, zowel vanuit kosteneffectiviteitsoverweging als om reeds gevaccineerde vrouwen gemotiveerd te houden voor deelname.
Toekomstig perspectief Medio 2019 heeft de Gezondheidsraad een update op haar eerdere advies uitgebracht.[44] Hierin wordt geadviseerd om naast meisjes ook jongens te vaccineren tegen HPV, om zodoende ook hun bescherming te bieden tegen de verschillende HPV-gerelateerde (voorloperstadia van) kanker. De ziektelast die kan worden veroorzaakt door een HPV-infectie bij mannen bestaat uit peniskanker, anuskanker en mond- en keelholtekanker. De vaccinatie zou aangeboden moeten worden rond het negende levensjaar, omdat is gebleken dat het vaccin langdurige bescherming biedt en om de kans dat er gevaccineerd wordt voor het seksueel debuut te vergroten. Daarnaast kan de mogelijkheid tot het combineren van de HPV-vaccinatie met de BMR- en DTP-vaccinatie die al op deze leeftijd worden aangeboden, leiden tot hogere deelname.
Volgens de Gezondheidsraad zal genderneutrale vaccinatie ervoor zorgen dat de transmissie van HPV in de populatie omlaag gaat en er grotere groepsbescherming mogelijk is, waarmee ook mannen die seks hebben met mannen (MSM) en ongevaccineerde meisjes beter beschermd zijn.[45] Tevens is de kosteneffectiviteitsratio van sekseneutrale vaccinatie zoals berekend onder de huidige omstandigheden, lager dan de gebruikte referentiewaarde van € 20.000 per Quality-Adjusted Life Year (QALY).[40] Dit gegeven gecombineerd met de veiligheid en effectiviteit van de vaccins maakt de belasting voor zowel het individu als het RVP als geheel aanvaardbaar, waarbij aanvaardbaarheid bepaald wordt door de verhouding tussen nut en risico van de vaccinatie. Ten slotte adviseerde de Gezondheidsraad om mensen tot en met 26 jaar de mogelijkheid te bieden zich alsnog te laten vaccineren tegen HPV via een aanvullend vaccinatieprogramma. In september 2019 werd duidelijk dat de minister van Volksgezondheid, Welzijn en Sport het advies van de Gezondheidsraad volgt; het streven is om vanaf 2021 te starten met het aangepaste programma.
Conclusie Tien jaar na de invoering van de HPV-vaccinatie blijkt uit vele nationale en internationale onderzoeken dat de effectiviteit van HPV-vaccinatie hoog en langdurig is en dat het vaccin veilig is. Hiermee heeft HPV-vaccinatie de potentie om een belangrijke rol te spelen in de preventie van verschillende HPV-gerelateerde kankers bij zowel mannen als vrouwen. De toenemende bewijzen voor effectiviteit tegen (voorloperstadia van) kanker en de geplande wijzigingen in het vaccinatieprogramma kunnen bijdragen aan het verbeteren van de vaccinatie-uptake en daarmee aan het verminderen van de totale HPV-gerelateerde ziektelast.
Syrjänen K, Hakama M, Saarikoski S, et al. Prevalence, incidence, and estimated life-time risk of cervical human papillomavirus infections in a nonselected Finnish female population. Sexually transmitted diseases. 1990;17:15-9.
De Villiers E-M, Fauquet C, Broker TR, Bernard H-U, zur Hausen H. Classification of papillomaviruses. Virology. 2004;324:17-27.
Baseman JG, Koutsky LA. The epidemiology of human papillomavirus infections. J Clin Virol. 2005;32:16-24.
de Martel C, Plummer M, Vignat J, Franceschi S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int J Cancer. 2017;141:664-70.
Doorbar J, Quint W, Banks L, et al. The biology and life-cycle of human papillomaviruses. Vaccine. 2012;30:F55-F70.
Gravitt PE. The known unknowns of HPV natural history. J Clin Invest. 2011;121:4593-9.
Moscicki A-B, Schiffman M, Burchell A, et al. Updating the natural history of human papillomavirus and anogenital cancers. Vaccine. 2012;30:F24-F33.
Ostör A. Natural history of cervical intraepithelial neoplasia: a critical review. Int J Gynecol Pathol. 1993;12:186-92.
Doorbar J. Molecular biology of human papillomavirus infection and cervical cancer. Clin Science. 2006;110:525-41.
Smith JS, Lindsay L, Hoots B, et al. Human papillomavirus type distribution in invasive cervical cancer and high‐grade cervical lesions: a meta‐analysis update. Int J Cancer. 2007;121:621-32.
Schurink-van 't Klooster TM, de Melker HE.
The National Immunisation Programme in the Netherlands: Surveillance and developments in 2017-2018. National Insitute for Public Health and the Environment; 2018. Report No.: 2018-0124.
Munoz N, Castellsagué X, de González AB, Gissmann L. HPV in the etiology of human cancer. Vaccine. 2006;24:S1-S10.
Bruni L, Diaz M, Castellsagué M, Ferrer E, Bosch FX, de Sanjosé S. Cervical human papillomavirus prevalence in 5 continents: meta-analysis of 1 million women with normal cytological findings. J Infect Dis. 2010;202:1789-99.
Kjaer SK, Chackerian B, Van Den Brule AJ, et al. High-risk human papillomavirus is sexually transmitted: evidence from a follow-up study of virgins starting sexual activity (intercourse). Cancer Epidemiology Prevention Biomarkers. 2001;10:101-6.
Wang JW, Roden RB. Virus-like particles for the prevention of human papillomavirus-associated malignancies. Expert review of vaccines. 2013;12:129-41.
Pedersen C, Petaja T, Strauss G, et al. Immunization of early adolescent females with human papillomavirus type 16 and 18 L1 virus-like particle vaccine containing AS04 adjuvant. Journal of Adolescent Health. 2007;40:564-71.
Reisinger KS, Block SL, Lazcano-Ponce E, et al. Safety and persistent immunogenicity of a quadrivalent human papillomavirus types 6, 11, 16, 18 L1 virus-like particle vaccine in preadolescents and adolescents: a randomized controlled trial. Pediatr Infect Dis J. 2007;26:201-9.
Muñoz N, Manalastas Jr R, Pitisuttithum P, et al. Safety, immunogenicity, and efficacy of quadrivalent human papillomavirus (types 6, 11, 16, 18) recombinant vaccine in women aged 24–45 years: a randomised, double-blind trial. The Lancet. 2009;373:1949-57.
Harper DM, Franco EL, Wheeler CM, et al. Sustained efficacy up to 4· 5 years of a bivalent L1 virus-like particle vaccine against human papillomavirus types 16 and 18: follow-up from a randomised control trial. The Lancet. 2006;36:1247-55.
Van Lier EA, Oomen P, van Vliet JA, Drijfhout IH, Zonnenberg-Hoff IF, de Melker HE. Vaccinatiegraad en jaarverslag Rijksvaccinatieprogramma Nederland 2018. National Institute for Public Health and the Environment; 2019. Report No.: 2019-0015.
Corcoran B, Clarke A, Barrett T. Rapid response to HPV vaccination crisis in Ireland. The Lancet. 2018;391:2103.
Schurink-van 't Klooster TM, van Kemmeren JM, Hilgersom WJA, Hoes J, de Melker HE. Surveillance van mogelijke bijwerkingen na HPV-vaccinatie: het vaccin is veilig Infectieziekten Bulletin. 2019;30(4).
Feiring B, Laake I, Bakken IJ, et al. HPV vaccination and risk of chronic fatigue syndrome/myalgic encephalomyelitis: a nationwide register-based study from Norway. Vaccine. 2017;35:4203-12.
Mollers M, King AJ, Knol MJ, et al. Effectiveness of human papillomavirus vaccine against incident and persistent infections among young girls: Results from a longitudinal Dutch cohort study. Vaccine. 2015;33:2678-83.
Donken R, King A, Bogaards J, Woestenberg P, Meijer C, de Melker H. High Effectiveness of the Bivalent Human Papillomavirus (HPV) Vaccine Against Incident and Persistent HPV Infections up to 6 Years After Vaccination in Young Dutch Women. J Infect Dis. 2018;217:1579-89.
van der Weele P, Breeuwsma M, Donken R, et al. Effect of the bivalent HPV vaccine on viral load of vaccine and non-vaccine HPV types in incident clearing and persistent infections in young Dutch females. PloS one. 2019;14:e0212927.
Woestenberg PJ, King AJ, van Benthem BHB, et al. Bivalent Vaccine Effectiveness Against Type-Specific HPV Positivity: Evidence for Cross-Protection Against Oncogenic Types Among Dutch STI Clinic Visitors. J Infect Dis. 2018;217:213-22.
Woestenberg PJ, Bogaards JA, King AJ, et al. Assessment of herd effects among women and heterosexual men after girls‐only HPV16/18 vaccination in the Netherlands: A repeated cross‐sectional study. Int J Cancer. 2019;144:2718-27.
Woestenberg PJ, King AJ, Van Benthem BHB, et al. Bivalent Vaccine Effectiveness Against Anal Human Papillomavirus Positivity Among Female Sexually Transmitted Infection Clinic Visitors in the Netherlands. J Infect Dis. 2019.
31. Bogaards JA, van der Weele P, Woestenberg PJ, van Benthem BH, King AJ. Bivalent HPV Vaccine Effectiveness Correlates with Phylogenetic Distance from Hpv Vaccine Types 16 And 18. The J Infect Dis. 2019. [epub ahead of print]
Drolet M, Bénard É, Pérez N, et al. Population-level impact and herd effects following the introduction of human papillomavirus vaccination programmes: updated systematic review and meta-analysis. The Lancet. 2019;394:497-509.
Pasmans H, Schurink-Van't Klooster TM, et al. Long-term HPV-specific immune response after one versus two and three doses of bivalent HPV vaccination in Dutch girls. Vaccine. 2019;37:7280-8.
van der Weele P, et al. Whole-genome sequencing and variant analysis of human papillomavirus 16 infections. J Virol. 2017;91:e00844-17.
Weele P, Meijer C, King A. High whole-genome sequence diversity of human papillomavirus type 18 isolates. Viruses. 2018;10:68.
van der Weele P, King AJ, Meijer CJ, Steenbergen RD. HPV16 variant analysis in primary and recurrent CIN2/3 lesions demonstrates presence of the same consensus variant. Papillomavirus Research. 2019;7:168-72.
Bogaards JA, Coupé VM, Xiridou M, Meijer CJ, Wallinga J, Berkhof J. Long-term impact of human papillomavirus vaccination on infection rates, cervical abnormalities, and cancer incidence. Epidemiology (Cambridge, Mass). 2011:505-15.
Matthijsse SM, et al. The estimated impact of natural immunity on the effectiveness of human papillomavirus vaccination. Vaccine. 2015;33:5357-64.
Bogaards JA, Kretzschmar M, Xiridou M, Meijer CJ, Berkhof J, Wallinga J. Sex-specific immunization for sexually transmitted infections such as human papillomavirus: insights from mathematical models. PLoS medicine. 2011;8:e1001147.
Qendri V, Bogaards JA, Berkhof J. Health and economic impact of a tender-based, sex-neutral human papillomavirus 16/18 vaccination program in the Netherlands. J Infect Dis. 2017;216:210-9.
Brisson M, Bénard É, Drolet M, et al. Population-level impact, herd immunity, and elimination after human papillomavirus vaccination: a systematic review and meta-analysis of predictions from transmission-dynamic models. The Lancet Public Health. 2016;1(1):e8-e17.
Palmer T, Wallace L, Pollock KG, et al. Prevalence of cervical disease at age 20 after immunisation with bivalent HPV vaccine at age 12-13 in Scotland: retrospective population study. BMJ. 2019;365:l1161.
Gezondheidsraad. advies Vaccinatie tegen HPV. 2019.
Brotherton JML, Giuliano AR, Markowitz LE, Dunne EF, Ogilvie GS. Monitoring the impact of HPV vaccine in males-Considerations and challenges. Papillomavirus research. 2016;2:106-11.
RIVM, Centrum voor Epidemiologie en Surveillance van Infectieziekten, Centrum voor Infectieziekten bestrijding (CIb), Bilthoven, J. Hoes, T.M. Schurink-van ’t Klooster, P.J. Woestenberg, B.H.B. van Benthem, J.A. Boogaards, H.E. de Melker. Amsterdam UMC, Vrije Universiteit, Amsterdam, afdeling Epidemiologie en Biostatistiek, J.A. Boogaards, J. Hoes. RIVM, Centrum voor Immunologie van Infectieziekten en Vaccins, Centrum voor Infectieziekten bestrijding (CIb), Bilthoven, H. Pasmans, F.R.M. van der Klis. Leids Universitair Medisch Centrum, Leiden, afdeling Medische Oncologie, H. Pasmans. RIVM, Centrum voor Infectieziekten Diagnostiek en laboratorium Surveillance, Centrum voor Infectieziekten bestrijding (CIb), Bilthoven, P. van der Weele, K. van Eer, A.J. King. Care and Public Health Research Institute (CAPHRI), Maastricht University Medical Center (MUMC+), Maastricht, P.J. Woestenberg. Correspondentieadres: J. Hoes (joske.hoes@rivm.nl).
<p><span class="Smaller90"><strong>Figuur 1.</strong> Incidentie van verschillende HPV-gerelateerde kankers in Nederland in 2018 per 100.000 en per leeftijdsgroep. Cijfers van www.cijfersoverkanker.nl (voorlopige data).</span></p>
<p><span class="Smaller90"><strong>Figuur 2.</strong> Vaccinatiegraad onder meisjes van verschillende geboortecohorten sinds invoering van de HPV-vaccinatie.</span></p>
<p><span class="Smaller90"><strong>Figuur 3.</strong> Vaccineffectiviteit tegen gecombineerde eindpunten (persisterende infecties) zeven jaar na vaccinatie op basis van de HAVANA-data.</span></p>
<p><span class="Smaller90"><strong>Figuur 4.</strong> Prevalentie van HPV16 en/of HPV18 bij vrouwen (A) en mannen (B) in de verschillende jaren waarin de PASSYON-studie is uitgevoerd, zowel voor als na invoering van vaccinatie (overgenomen in aangepaste vorm van Woestenberg et al.).[27]</span></p>
<p><span class="Smaller90"><strong>Figuur 5.</strong> Geometrisch gemiddelde antistofconcentratie tegen verschillende HPV-typen bij deelneemsters aan de HPV2D-studie (gevaccineerd met twee doses) tot en met 48 maanden na vaccinatie (voorlopige resultaten).</span></p>
<p> </p>
<p>Bestrijding van ebola anno 2019<br /><span class="Smaller90">De rol van beschikbare vaccinaties en behandelingen</span></p>
Laura Doornekamp, Eric van Gorp, Marion Koopmans
Thema: Vaccinaties (2)
2019;27:nr4
Ebolavirus, epidemie, uitbraak, volksgezondheid
Samenvatting De huidige ebola-uitbraak in Democratische Republiek Congo (DR Congo) is, na die in West-Afrika, de grootste wereldwijd. Door de epidemie in West-Afrika in 2013-2016 is onderzoek naar vaccins en behandelingen voor ebola in een stroomversnelling gekomen. Destijds waren er niet genoeg middelen beschikbaar om direct in te zetten en om klinisch te onderzoeken. Om in de toekomst beter voorbereid te zijn, heeft de WHO de R&D blueprint for emerging infections opgericht. Hiermee wordt onder andere gestreefd wetenschappelijk onderzoek naar (genees)middelen versneld op te zetten. Dit gebeurde tijdens de uitbraak in DR Congo die in augustus 2018 is ontstaan: het levend verzwakte vaccin rVSV-ZEBOV-GP is vanaf het begin op grote schaal ingezet en in het najaar van 2019 is gestart met het toedienen van het experimentele combinatievaccin Ad26.ZEBOV en MVA-BN-Filo. Daarnaast is er vroeg in de uitbraak een grote studie gestart waarin de effectiviteit van vier geneesmiddelen is getest. Uit de interim-analyse is geconcludeerd dat REGN-EB3 en mAb114 grotere overlevingskansen geven ten opzichte van de andere twee middelen. De uitkomsten van dit soort onderzoeken naar preventieve en therapeutische mogelijkheden zijn veelbelovend. De krachten van hulpverleners, politici, geldschieters en de lokale bevolking moeten echter worden gebundeld om de epidemie onder controle te krijgen.
Abstract The current Ebola outbreak in Democratic Republic of the Congo (DR Congo) is the second largest worldwide, after the one in West-Africa. The epidemic in West-Africa in 2013-2016 boosted research into the development of vaccines and therapies for Ebola. At that time, vaccines and medicines were unavailable for phase 2 or 3 trials. To be better prepared in the future, the WHO launched the R&D blueprint for emerging infections. The aim of this blueprint is to accelerate the availability of vaccines and medicines for emerging, threatening pathogens. Early after the onset of the Ebola outbreak in DR Congo in August 2018 the rVSV-ZEBOV-GP vaccine was introduced. The experimental use of the combination vaccine Ad26.ZEBOV and MVA-BN-Filo was started in the autumn of 2019. Additionally, a large trial was launched early in the outbreak to investigate the effectiveness of four medicines in the treatment for Ebola. The interim-analysis showed that REGN-EB3 and mAb114 had better survival rates compared to the other two therapies. The results of vaccine and treatment trials like these are promising. However, frontline workers, politicians, funders and locals have to combine their forces to combat the epidemic.
Ebola-uitbraak DR Congo anno 2019 Democratische Republiek Congo (DR Congo) maakt voor de tiende maal een ebola-epidemie door, de grootste die het land ooit heeft gekend. Na de snelle indamming van de negende uitbraak in de Evenaarsprovincie van dit land in mei 2018, had men gehoopt dat ook deze uitbraak, die begin augustus 2018 startte, beperkt zou blijven. Deze ebola-uitbraak ontstond echter in Noord-Kivu, een provincie in het noordoosten van het land. In dit deel van het land iseen humanitaire crisis gaande, wat de bestrijding veel lastiger maakt. Bijna een jaar later, op 17 juli 2019, kondigde de WHO een ‘Public Health Emergency of International Concern’ (PHEIC) af, vanwege de verspreiding naar de miljoenenstad Goma. Met ruim 3000 ebolagevallen en meer dan 2000 overledenen eind augustus 2019 is de huidige uitbraak de op één na grootste ooit wereldwijd.[1] Ten opzichte van de grootste uitbraak allertijden, in West-Afrika van 2013 tot 2016, zijn er nu echter meer kennis en middelen beschikbaar om de uitbraak te bestrijden. In vervolg op een artikel in het Tijdschrift voor Infectieziekten bespreken we in dit artikel de status van beschikbare vaccins en behandelingen.[2]
Ebolavirus – de boosdoener Ebolavirus behoort samen met Marburgvirus tot de familie van Filoviridae. Het ebolavirus dat het meest frequent de kop op steekt in Afrika is het Zaïreebolavirus. De andere subtypen die momenteel bekend zijn, zijn Sudan, Reston, Taï Forest en Bundibugyo, en het recent gekarakteriseerde Bombali-virus.[3,4] Van vier van de zes subtypen is bekend dat ze ziekte kunnen veroorzaken bij mensen. Het Restonebolavirus is niet humaan pathogeen en hoewel nieuwe data laten zien dat het Bombaliebolavirus humane cellen kan infecteren, is nog niet bekend of het virus ook ziekte veroorzaakt bij mensen.[4].
Het ziektebeeld dat veroorzaakt wordt door het Zaïre ebolavirus was voorheen bekend onder de naam ‘Ebola Hemorrhagic Fever’, maar is om verwarring te voorkomen veranderd naar Ebola Virus Disease omdat de infectie slechts in vijf procent van de gevallen gepaard gaat met significante klinische bloedingen.[5] Een infectie met het Zaïre ebolavirus geeft voornamelijk koorts, maculopapulaire huiduitslag en griepachtige klachten, maar ook gastro-intestinale, respiratoire, neurologische en oculaire symptomen komen frequent voor. Later in het beloop van de ziekte treedt niet zelden orgaanfalen op, maar bij degenen die overleven worden dan cellulaire en humorale responses waargenomen.[5] De incubatietijd varieert van twee tot 21 dagen en overlevenden kunnen nog lang besmettelijk blijven.[3] Viraal RNA kan vanaf het begin van de klachten worden aangetoond in het bloed via een kwantitatieve real-time polymerase chain reaction (qRT-PCR), met een piek in de virale load een ruime week na het begin van ziekte. Het risico op mens-tot-menstransmissie is het grootste bij direct contact met lichaamsvloeistoffen van een symptomatische patiënt. Gezondheidswerkers lopen hierdoor een hoog risico op besmetting. Ook na de dood is het lichaam nog besmettelijk: infectieus virus is geïsoleerd uit kadavers tot negen dagen na overlijden. Dit vraagt om beschermingsmaatregelen bij begrafenissen en bijbehorende rituelen.[6]
De ebola-uitbraak in West-Afrika heeft enkele opvlammingen gehad door seksuele transmissie. Ebolavirus-RNA is aangetoond in het sperma van Liberiaanse overlevenden tot ruim 18 maanden na herstel.[7] Of dit echter infectieus virus is, is onduidelijk. Wel zijn er casussen beschreven die seksuele transmissie van mannelijke overlevenden naar vrouwelijke partners, na 179 dagen, waarschijnlijk hebben gemaakt via genoomsequentie.[5] Vanwege deze bevindingen heeft de WHO mannelijke overlevenden geadviseerd om tot 12 maanden na herstel of tot tweemaal negatief getest sperma geen onbeschermd seksueel contact te hebben. Hoewel dit advies moeilijk navolgbaar lijkt, rapporteert 76 procent van de mannelijke overlevenden in dezelfde Liberiaanse vervolgstudie condoomgebruik bij hun laatste seksuele contact voordat hun sperma tweemaal negatief is getest. Dit onderstreept het belang van deze studies en gedragsadviezen.[7]
Gestelde doelen voor ebolavaccins Bovenstaande illustreert dat er verschillende situaties zijn waarbij vaccins ingezet kunnen worden. Deze situaties en bijbehorende karakteristieken voor de benodigde vaccins zijn beschreven in de Research & Development (R&D) blueprint for emerging infections, opgezet door de WHO naar aanleiding van de ebola-epidemie in West-Afrika.[8] De R&D blueprint bestaat uit drie pijlers: (1) het verbeteren van de coördinatie van de uitbraakrespons; (2) het versnellen van wetenschappelijk onderzoek naar (genees)middelen; (3) het ontwikkelen van normen en standaarden voor het uitvoeren van klinische studies en het delen van monsters en data. In het artikel in het Tijdschrift voor Infectieziekten gepubliceerd in februari jongstleden wordt voornamelijk de eerste pijler toegelicht.[2] Hier zullen we verder ingaan op de tweede en derde pijler. De tweede pijler focust op R&D. De WHO heeft een prioriteitenlijst opgesteld voor pathogenen die mogelijk een epidemie met grote impact zouden kunnen veroorzaken. Voor deze infectieziekten worden zogeheten ‘roadmaps’ en ‘target product profiles (TPP)’ opgesteld. De roadmaps bevatten korte overzichten van de belangrijkste beschikbare kennis over het virus en de kennishiaten waarnaar meer onderzoek nodig is. TPP’s beschrijven de karakteristieken van de te ontwikkelen en te testen diagnostische methoden, vaccins en geneesmiddelen.[8] De TPP voor ebola is verdeeld in twee scenario’s: één voor reactief gebruik en één voor profylactisch gebruik. Met reactief gebruik wordt met name gerefereerd aan vaccins die worden ingezet tijdens een uitbraak om verdere verspreiding in de gemeenschap te voorkomen, terwijl profylactische vaccinatie met name hulpverleners en personeel van zorginstellingen in risicogebieden betreft. Beide scenario’s vragen om andere karakteristieken van vaccins, die worden beschreven in de TPP (tabel 1).[9] Voordat een vaccin geproduceerd wordt, moet het geregistreerd worden bij een nationale regelgevende autoriteit en moet een WHO-prekwalificatie in gang gezet worden. In dit prekwalificatieproces wordt gekeken naar kwaliteit, veiligheid, werkzaamheid en geschiktheid van het vaccin, specifiek voor laag- en middeninkomenslanden.[9]
De derde pijler omvat het opzetten van klinische studies en het delen van data en monsters via open platforms. In het algemeen worden na de ontdekking van potentieel werkzame middelen veiligheid, toxiciteit en farmacodynamiek en -kinetiek getest via in-vitrostudies. Als deze kandidaatmiddelen zich in preklinische fasen hebben bewezen, kunnen ze bij mensen getest worden. Deze klinische studies worden ingedeeld in drie fasen. In de eerste fase, die vaak enkele maanden duurt, wordt het middel aan maximaal enkele tientallen gezonde mensen toegediend en worden de veiligheid en dosering onderzocht. Tijdens fase 2-studies worden verdere veiligheid, werkzaamheid (in geval van vaccins vaak via immuunrespons) en voorspelbare bijwerkingen onderzocht bij honderden mensen met potentiële blootstelling. Dit kan enkele maanden tot jaren duren, maar als het middel goede uitkomsten heeft kan het door naar een fase 3-studie, waar werkzaamheid en mogelijk schadelijke bijwerkingen centraal staan. Honderden tot duizenden individuen in een setting waar ziekte voorkomt krijgen het middel toegediend en de studie kan een aantal jaren duren. Tot slot wordt van een fase 4-studie gesproken als het middel op de markt is gekomen, maar blijvend wordt geëvalueerd.[10]
Voor opkomende virusinfecties is het uitvoeren van fase 2- en 3-studies vaak lastig omdat (potentiële) patiënten nodig zijn om de werkzaamheid van een vaccin te testen. Wanneer een uitbraak plaatsvindt, zouden er dus middelen op de plank moeten liggen die succesvol uit een fase 1-studie zijn gekomen. Dat was niet het geval ten tijde van de omvangrijke ebola-uitbraak in West-Afrika.[11] In januari 2016 is hier met de oprichting van de Coalition for Epidemic Preparedness Innovations (CEPI) hopelijk verandering in gekomen. CEPI werft financiële middelen om versneld vaccins te ontwikkelen voor ziekten op de prioriteitenlijst van de WHO, waarbij het accent ligt op klinische studies.[12]
Stand van zaken vaccins Tot 2014 waren slechts twee ebolavirusvaccins getest in fase 1-klinische studie, één gebaseerd op een DNA-platform en de ander op een adenovirusvector.[13] De uitbraak in West-Afrika bracht vaccinontwikkeling in een stroomversnelling: een vaccin voor ebola was hard nodig en de mogelijkheden voor fase 2- en 3-studies boden zich aan. Van de vaccins die verder dan fase 1 onderzocht waren, was echter onvoldoende voorraad om studies snel te starten en preklinische data waren beperkt beschikbaar.[11] Na de afkondiging van de PHEIC in augustus 2014 zijn de inspanningen om vaccins beschikbaar te maken bespoedigd en werden er snel twee vaccins geïntroduceerd die onderzocht werden in fase 1-studies. Een van de twee was een replicatiecompetent vectorvaccin, genaamd rVSV-ZEBOV-GP, en de andere een replicatie-incompetent vectorvaccin, ChAd3-EBOZ. rVSV-ZEBOV-GP is een recombinant vesiculair stomatitis virus dat een glycoproteïne (GP) van het Zaïre-subtype tot expressie brengt en dus in staat is om zich te repliceren. De resultaten van een gecombineerde fase 1/2-studie met dit vaccin waren positief en een fase 3-studie kon dus gestart worden. In april 2015 is een cluster gerandomiseerd onderzoek (genaamd Ebola Ça Suffit!) gestart in Guinee, en later ook in Sierra Leone. Clusters van contacten van cases en contacten van deze contacten zijn gerandomiseerd in groepen die direct vaccinatie ontvingen en groepen die na 21 dagen vaccinatie ontvingen. Een interimanalyse na vier maanden liet een geschatte effectiviteit van 100 procent zien, waarbij – mede onder druk van de omstandigheden – werd besloten om de randomisatie stop te zetten en iedereen direct te vaccineren. In totaal zijn ruim 5800 personen gevaccineerd, inclusief bijna 200 kinderen van 6 tot 18 jaar, waarbij geen ernstige bijwerkingen zijn opgetreden, met goede resultaten wat betreft effectiviteit en werkzaamheid.[14] Later zijn echter de conclusies over bescherming betwist, de vergelijkbaarheid van alle variabelen behalve de vaccinatie voor de twee studiegroepen werd bediscussieerd.[15] Daarnaast is de duur en mate van immuunrespons in deze studie niet bestudeerd.
Het andere kandidaat vaccin, ChAd3-EBOZ – dat chimpansee adenovirustype 3 als backbone heeft en zich niet kan repliceren – was pas klaar voor een fase 3-studie toen de epidemie al vrijwel onder controle was.[16] Daardoor is de nadruk in de gecombineerde fase 2/3-studie (genaamd PREVAIL) die plaatsvond in Liberia, meer op fase 2 komen te liggen. In deze studie zijn 1500 deelnemers gerandomiseerd voor vaccinatie met rVSV-ZEBOV-GP, ChAd3-EBOZ of placebo en werden IgG-antistofresponsen bestudeerd. Hoewel 1 week post-vaccinatie slechts 2,5 procent van de rVSV-ZEBOV-GP-ontvangers en 3,8 procent van de ChAd3-EBOZ-ontvangers was geseroconverteerd, was na een maand respectievelijk 84 en 71 procent en na 12 maanden respectievelijk 80 en 64 procent geseroconverteerd.[17] Andere studies hebben de cellulaire immuunrespons voor rVSV-ZEBOV-GP bestudeerd en laten zien dat dit vaccin ook T-celimmuniteit genereert en handhaaft tot 56 dagen na een enkele vaccinatie (met de dosis gebruikt in de Ebola Ça Suffit!-studie). Een ‘correlate of protection’ is echter nog niet vastgesteld voor ebola, dus de mate van bescherming is lastig te interpreteren.[18] De verklaring voor het feit dat ChAd3-EBOZ een lagere immuunrespons gaf, wordt gezocht in de reeds bestaande immuniteit voor adenovirus 3 onder grote delen van de bevolking.[16] Later werden daarom andere adenovirussen ingezet, humaan en met een lagere prevalentie van immunologisch geheugen, zoals in de ebolavaccins Ad5-EBOV en Ad26. Figuur 1 laat zien in welke onderzoeksfasen deze vaccins zich bevinden, als homoloog vaccin of als onderdeel van een prime-boostserie.[19]
Andere vaccins in ontwikkeling zijn Modified Vaccinia Ankara (MVA)-vaccins en DNA-vaccins. Deze typen vaccins worden als toepassing voor verschillende infectieziekten onderzocht. MVA-vaccins bestaan uit replicatie-incompetente, verzwakte vectoren die zijn ontwikkeld uit vacciniavirus en werken door antigenen tot expressie te brengen in gastheercellen tijdens infectie. DNA-vaccins worden gezien als een veelbelovend platform voor de toekomst, omdat ze gemakkelijk en voordelig te produceren zijn. Bij intramusculaire toediening zijn ze echter laag-immunogeen omdat het DNA in de celkern moet komen. Elektroporatie - dat wil zeggen het opwekken van een elektrische puls op de huid - is dan succesvoller. Elektroporatie zorgt voor een verstoring van het celmembraan, waardoor het vaccin op de juiste plek komt en er effectieve antigenproductie optreedt.[20] Het belangrijkste MVA-kandidaatvaccin voor ebola is MVA-BN-Filo. Dit vaccin wordt gebruikt voor een heteroloog prime-booster vaccinatieschema in combinatie met Ad26.ZEBOV. Uit de fase 1-studies, geïnitieerd in Engeland eind 2014 en later in onder andere in Kenia, Oeganda en Tanzania, werd geconcludeerd dat de vaccins goed getolereerd werden en immunogeen waren.[20,21] Data van een fase 3-studie in Sierra Leone zullen naar verwachting binnenkort beschikbaar komen.[22] Een ander voordeel van deze vaccincombinatie, is dat het lijkt te voldoen aan de gewenste stabiliteit- en opslagcapaciteiten, iets dat bij het rVSV-ZEBOV-GP-vaccin nog als een bemoeilijkende factor wordt gezien.[23] Gezien de voortreffelijke uitkomsten van het rVSV-ZEBOV-GP-vaccin op gebied van effectiviteit, zijn tijdens de twee latere ebola-uitbraken in DR Congo spoed-vaccinatiecampagnes met alleen dit vaccin gestart. Ondertussen zijn meer dan 222.000 mensen gevaccineerd met het rVSV-ZEBOV-GP-vaccin, onder wie 46.000 gezondheids- en frontliniewerkers, ruim 73.000 kinderen (van wie 1160 jonger dan 1 jaar), 800 zwangere vrouwen en meer dan 4000 moeders die borstvoeding geven.[24]
Om ervoor te zorgen dat er voorlopig voldoende vaccins beschikbaar blijven, is recent de dosis van rVSV-ZEBOV-GP teruggebracht van 1 ml naar 0,5 ml (zoals effectief bleek in de Ebola Ça Suffit!-studie) of 0,2 ml voor groepen met een laag risico op infectie.[25] Daarnaast heeft het ministerie van gezondheid van DR Congo eind juni 2019 aangegeven open te staan voor het starten van studies met andere vaccins.[26] Als antwoord hierop werd op 23 september 2019 aangekondigd dat een nieuwe studie gestart wordt met de vaccincombinatie Ad26.ZEBOV en MVA-BN-Filo (met 56 dagen interval tussen de primovaccinatie en de booster) met gerichte geografische en pop-up vaccinatiecampagnes.[27] Op 12 november 2019 is het rVSV-ZEBOV-GP-vaccin goedgekeurd door de WHO, na een positief advies van de Europese Commissie en het Europese Medicijn Agentschap (EMA). Het vaccin wordt op de markt gebracht onder de naam Ervebo.[28]
Stand van zaken behandelingen Ondertussen zijn er ook verschillende kandidaattherapieën voor ebola op de markt, welke lijken bij te dragen aan verbeterde overlevingskansen. Drie geneesmiddelen, ZMapp, favipiravir en remdesivir (ook wel GS-5734 genoemd), zijn reeds experimenteel gebruikt tijdens de epidemie in West-Afrika. Doordat onderzoek pas laat op gang kwam, zijn deze middelen slechts op kleine schaal getest en is de werkzaamheid van geen van de middelen nog bewezen.[29] Toen vervolgens ebola uitbrak in de Evenaarsprovincie van DR Congo, is voor het gebruik van vijf experimentele therapieën toestemming gegeven onder het WHO ethisch raamwerk. Deze uitbraak werd echter spoedig ingedamd. De vijf experimentele middelen - ZMapp, favipiravir, remdesivir, mAb114 en REGN-EB3 (ook wel Regn3470-3471-3479 genoemd) - konden vervolgens alsnog snel ingezet worden toen ebola uitbrak in Noord-Kivu.[30]
Over het algemeen worden antivirale middelen ingedeeld in direct werkende, indirect werkende en gastheergerichte therapieën. Favipiravir en remdesivir zijn beide direct werkende antivirale middelen. Ze vallen onder de nucleoside analogen, die aangrijpen op de RNA-synthese van ebolavirus. De werking van de andere drie genoemde middelen is gebaseerd op monoklonale antilichamen die glycoproteïnen van ebolavirus binden. ZMapp is een fusie-inhibitor die het intredeproces van ebolavirus remt. mAb114 is één monoklonaal antilichaam die sterke neutraliserende capaciteiten heeft door te binden aan de oppervlakte-eiwitten van het virus. REGN-EB3 bevat drie monoklonale antilichamen met drie verschillende aangrijpingspunten.[31]
Op 24 november 2018 is een gerandomiseerde, gecontroleerde fase 2/3-studiie (de PALM-studie) gestart in DR Congo, waarbij vier verschillende behandelingen werden vergeleken: ZMapp, remdesivir, mAb114 en REGN-EB3. Na ruim acht maanden, op 9 augustus 2019, werd tijdens een interimanalyse van de behandelingen van bijna 500 patiënten geconcludeerd dat REGN-EB3 en mAb114 grotere overlevingskansen geven ten opzichte van de andere twee middelen.[24] Wanneer de patiënt zich tijdens de vroege fase van de infectie presenteerde, was de mortaliteit onder patiënten behandeld met remdesivir 33 procent, met ZMapp 24 procent, 11 procent met mAb114 en 6 procent met REGN-EB3. In de praktijk presenteren patiënten zich vaak pas later in het ziekteproces in een gezondheidscentrum, zeker in de regio van de huidige uitbraak. Onder die omstandigheden waren de voorlopige resultaten minder gunstig, met mortaliteitscijfers van respectievelijk 53, 49, 34 en 29 procent. Aan de hand van deze resultaten is toch besloten de studie voort te zetten met alleen REGN-EB3 en mAb114 en de gegevens van de interimanalyse te gebruiken om personen met klachten na contact met een ebolapatiënt, te adviseren zich zo snel mogelijk te melden. Naar verwachting zullen in oktober 2019 meer data beschikbaar komen, aangezien de totale studie was berekend op circa 750 personen.[32]
Daarnaast blijft het handhaven van de vocht- en elektrolytenbalans en symptoombestrijding van groot belang. Dit blijkt uit de relatief lage mortaliteit (18,5 procent) van de patiënten behandeld met goede ondersteunende zorg, in sommige gevallen ook aangevuld door experimentele behandelingen, in Westerse landen.[5]
Uitdagingen in vaccinatie- en behandelstrategieën De voorzichtig positieve resultaten uit klinische studies voor zowel vaccins als behandelstrategieën zullen eraan moeten bijdragen dat de lokale bevolking vertrouwen krijgt in autoriteiten en hulpverleningsorganisaties, zodat de perceptie verandert dat iedereen die naar een ebolabehandelcentrum gaat, overlijdt.[33] Naast dit wantrouwen heeft de huidige epidemie in DR Congo een andere factor die de uitbraakbestrijding zeer bemoeilijkt: de humanitaire crisis die zich in de dichtbevolkte regio van het land afspeelt, bedreigt de veiligheid van hulpverleners en belemmert het opsporen van contacten door ontheemde of gevluchte families.[30]
Toekomstblik Hoewel de uitkomsten van onderzoek naar preventieve en therapeutische mogelijkheden veelbelovend lijken, is de ebola-uitbraak in DR Congo nog niet ten einde. Naast het doorzetten en uitbreiden van medisch-wetenschappelijk onderzoek, onder andere naar de duur van immuniteit, effectiviteit van vaccinatie op chronisch dragerschap, aanvullende vaccins met bredere en langdurige bescherming en optimale opslagomstandigheden, is ook antropologisch onderzoek dat kan bijdragen aan succesvolle uitbraakbestrijding, van groot belang. Als men beter inzicht heeft in de kennis, attitude en het gedrag van de lokale bevolking, dan hebben voorlichtings- en interventiecampagnes meer kans van slagen. Wanneer niet alleen hulpverleners, politici en geldschieters, maar ook de mensen die door de uitbraak worden getroffen, hun krachten bundelen, neemt de kans toe dat we met de huidige middelen de epidemie spoedig onder controle krijgen.
MSF. Crisis Update - August 2019: Medicins Sans Frontieres; 2019 [updated 13 september 2019]. Available from: https://www.msf.org/drc-ebola-outbreak-crisis-update.
Doornekamp L, van Gorp ECM, Koopmans MPG. De strijd tegen ebola - lering getrokken uit de epidemie in West-Afrika. Tijdschr Infect. 2019;14:11-7.
Goeijenbier M, van Kampen JJ, Reusken CB, et al. Ebola virus disease: a review on epidemiology, symptoms, treatment and pathogenesis. Neth J Med. 2014;72:442-8.
Goldstein T, Anthony SJ, Gbakima A, et al. The discovery of Bombali virus adds further support for bats as hosts of ebolaviruses. Nat Microbiol. 2018;3:1084-9.
Baseler L, Chertow DS, Johnson KM, et al. The Pathogenesis of Ebola Virus Disease. Annual Review of Pathology: Mechanisms of Disease. 2017;12:387-418.
Vetter P, Fischer WA, 2nd, Schibler M, et al. Ebola Virus Shedding and Transmission: Review of Current Evidence. J Infect Dis. 2016;214:S177-S84.
Soka MJ, Choi MJ, Baller A, et al. Prevention of sexual transmission of Ebola in Liberia through a national semen testing and counselling programme for survivors: an analysis of Ebola virus RNA results and behavioural data. Lancet Glob Health. 2016;4:e736-43.
WHO. R&D Blueprint 2015. Available from: http://www.who.int/blueprint/what/en/.
WHO. WHO Ebola Vaccine Target Product Profile. 2016 5 januari 2016. Report No.
10. FDA. Clinical Research: U.S. Food & Drug Administration; 2018 [updated 1 april 2018. Available from: https://www.fda.gov/patients/drug-development-process/step-3-clinical-research#Clinical_Research_Phase_Studies.
Plotkin SA, Mahmoud AAF, Farrar J. Establishing a Global Vaccine-Development Fund. N Engl J Med. 2015;373:297-300.
CEPI. CEPI wants to galvanise the development of new vaccines against diseases we know could cause the next devastating epidemic. 2017. Available from: http://cepi.net/approach#What-CEPI-will-do-thats-different.
Ledgerwood JE, Costner P, Desai N, et al. A replication defective recombinant Ad5 vaccine expressing Ebola virus GP is safe and immunogenic in healthy adults. Vaccine. 2010;29:304-13.
Henao-Restrepo AM, Camacho A, Longini IM, et al. Efficacy and effectiveness of an rVSV-vectored vaccine in preventing Ebola virus disease: final results from the Guinea ring vaccination, open-label, cluster-randomised trial (Ebola Ca Suffit!). The Lancet. 2017;389:505-18.
Metzger WG, Vivas-Martinez S. Questionable efficacy of the rVSV-ZEBOV Ebola vaccine. Lancet. 2018;391:1021.
Venkatraman N, Silman D, Folegatti PM, et al. Vaccines against Ebola virus. Vaccine. 2018; 28;36:5454-9.
Kennedy SB, Bolay F, Kieh M, et al. Phase 2 Placebo-Controlled Trial of Two Vaccines to Prevent Ebola in Liberia. N Engl J Med. 2017;377:1438-47.
Farooq F, Beck K, Paolino KM, et al. Circulating follicular T helper cells and cytokine profile in humans following vaccination with the rVSV-ZEBOV Ebola vaccine. Scientific Reports. 2016;6:27944.
WHO. R&D Blueprint Mapping Tool: WHO; 2019. Available from: http://who-blueprint-mapping-tool.surge.sh/.
Martins KA, Jahrling PB, Bavari S, et al. Ebola virus disease candidate vaccines under evaluation in clinical trials. Expert Review of Vaccines. 2016;15:1101-12.
Anywaine Z, Whitworth H, Kaleebu P, et al. Safety and Immunogenicity of a 2-Dose Heterologous Vaccination Regimen With Ad26.ZEBOV and MVA-BN-Filo Ebola Vaccines: 12-Month Data From a Phase 1 Randomized Clinical Trial in Uganda and Tanzania. J Infect Dis. 2019;220:46-56.
Janssen. Staged Phase 3 Study to Assess the Safety and Immunogenicity of Ebola Candidate Vaccines Ad26.ZEBOV and MVA-BN-Filo During Implementation of Stages 1 and 2 (EBOVAC-Salone): ClinicalTrials.gov; 2019 [updated 18 juli 2019. Available from: https://clinicaltrials.gov/ct2/show/NCT02509494.
Capelle MAH, Babich L, van Deventer-Troost JPE, et al. Stability and suitability for storage and distribution of Ad26.ZEBOV/MVA-BN(R)-Filo heterologous prime-boost Ebola vaccine. Eur J Pharm Biopharm. 2018;129:215-21.
WHO. Ebola virus disease - Democratic Republic of the Congo: External Situation Report 59: WHO; 2019 [updated 17 september 2019. Available from: https://www.who.int/publications-detail/ebola-virus-disease-democratic-republic-of-congo-external-situation-report-59-2019.
WHO. WHO adapts Ebola vaccination strategy in the Democratic Republic of the Congo to account for insecurity and community feedback: WHO; 2019 [updated 7 mei 2019. Available from: https://www.who.int/news-room/detail/07-05-2019-who-adapts-ebola-vaccination-strategy-in-the-democratic-republic-of-the-congo-to-account-for-insecurity-and-community-feedback.
Nakkazi E. Apply to trial Ebola vaccines in DR Congo, says ministry. The Lancet. 2019;394:15-6.
WHO. Second Ebola vaccine to complement “ring vaccination” given green light in DRC: WHO; 2019 [updated 23 september 2019. Available from: https://www.who.int/news-room/detail/23-09-2019-second-ebola-vaccine-to-complement-ring-vaccination-given-green-light-in-drc.
WHO. WHO prequalifies Ebola vaccine, paving the way for its use in high-risk countries: WHO; 2019 [updated 12 November 2019. Available from: https://www.who.int/news-room/detail/12-11-2019-who-prequalifies-ebola-vaccine-paving-the-way-for-its-use-in-high-risk-countries.
Hayden EC. Experimental drugs poised for use in Ebola outbreak: Nature; 2018 [updated 18 May 2018. Available from: https://www.nature.com/articles/d41586-018-05205-x.
WHO. Ebola virus disease - Democratic Republic of the Congo: External Situation Report 03: WHO; 2018 [updated 22-08-2018. Available from: http://apps.who.int/iris/bitstream/handle/10665/274258/SITREP_EVD_DRC_20180820-eng.pdf?ua=1.
Hoenen T, Groseth A, Feldmann H. Therapeutic strategies to target the Ebola virus life cycle. Nat Rev Microbiol. 2019;17:593-606.
Dyer O. Two Ebola treatments halve deaths in trial in DRC outbreak. BMJ. 2019;366:l5140.
WHO. Factors that contributed to undetected spread of the Ebola virus and impeded rapid containment: WHO; 2015. Available from: http://www.who.int/csr/disease/ebola/one-year-report/factors/en/.
Erasmus Medisch Centrum, afdeling Viroscience, Rotterdam, drs. L. Doornekamp, arts-onderzoeker, vaccinatie-arts, TravelClinic. Erasmus Medisch Centrum, Rotterdam, afdeling Interne Geneeskunde, dr. E.C.M. van Gorp, viroloog afdeling Viroscience. Erasmus Medisch Centrum, Rotterdam, afdeling Viroscience, prof. dr. Marion Koopmans, dierenarts-viroloog. Correspondentieadres: prof. dr. Marion Koopmans, m.koopmans@erasmusmc.nl
<p><span class="Smaller90"><strong>Tabel 1. </strong>Vereiste en gewenste karakteristieken voor ebolavaccins voor reactief en profylactisch gebruik[9]</span></p>
Reactief gebruik
Profylactisch gebruik
Type vaccin
Monovalent (tegen Zaïre ebolavirus) is voldoende, bredere bescherming tegen eveneens Sudan ebolavirus, Marburg virus, Bundibugyo ebolavirus gewenst.
Voorkeur voor bredere dekking, naast Zaïre ebolavirus, ook bescherming tegen Sudan ebolavirus, Marburg virus en Bundibugyo ebolavirus gewenst.
Doelgroep
Ten minste alle volwassenen, maar liever nog alle leeftijdscategorieën inclusief bijzondere populaties (zoals zwangeren).
Tenminste alle volwassenen, maar liever nog alle leeftijdscategorieën inclusief bijzondere populaties (zoals zwangeren).
Vaccinatieschema
Grote voorkeur voor eenmalige vaccinatie. Zeker niet meer dan twee doses, met maximaal één maand interval, waarbij na eerste dosis al enige mate van bescherming moet zijn opgebouwd. Indien twee doses, dan met homologe vaccins.
Voorkeur voor eenmalige vaccinatie. Primaire serie: niet meer dan twee doses, maximaal één maand interval. Voorkeur homoloog vaccin, maar heteroloog prime-boost serie optioneel. Boosterdoses niet meer dan jaarlijks of ten tijde van uitbraak.
Effectiviteit
Bij voorkeur ten minste 80 procent, maar ten minste 50 procent bij gezonde volwassenen.
Bij voorkeur ten minste 80 procent, maar ten minste 70 procent bij gezonde volwassenen.
Tijd tot bescherming
Nadruk op snelle bescherming (bij voorkeur binnen twee weken, ten minste binnen één maand).
Nadruk meer op langdurige bescherming. Tijd tot bescherming niet gedefinieerd.
Beschermingsduur
Ten minste drie maanden bescherming, maar bij voorkeur één jaar.
Minimaal één jaar bescherming na primaire serie, maar bij voorkeur minimaal vijf jaar. Kan onderhouden worden met boosters.
Toediening
Injecteerbaar (IM, ID of SC), oraal of niet-parenteraal alternatief gewenst.
Injecteerbaar (IM, ID of SC), oraal of niet-parenteraal alternatief gewenst.
Kan samen met andere vaccins toegediend worden, indien bestemd voor dezelfde populaties.
Verpakking
Als mono- of multidoses verpakking aan te leveren, maximaal doseringsvolume van 0,5 ml. Als vloeistof of gevriesdroogd met gepast oplosmiddel.
Als mono- of multidoses verpakking aan te leveren, maximaal doseringsvolume van 0,5 ml. Als vloeistof of gevriesdroogd met gepast oplosmiddel.
Stabiliteit en opslag
Hoog. Moet minimaal 12 maanden op -80⁰C kunnen worden opgeslagen en stabiel zijn voor minimaal acht uur op 2-8⁰C. Grotere voorkeur heeft nog een houdbaarheid van minimaal 24 maanden op -20⁰C en zes maanden stabiliteit op 2-8⁰C.
Minder van belang. Moet minimaal 12 maanden op -20⁰C kunnen worden opgeslagen en stabiel zijn voor minimaal acht uur op 2-8⁰C. Grotere voorkeur heeft nog een houdbaarheid van minimaal 24 maanden op -20⁰C en zes maanden stabiliteit op 2-8⁰C.
Productiecapaciteit
Capaciteit van ten minste 10.000 doses per maand met mogelijkheid tot direct vergroten productie. Bij voorkeur minimaal vijf miljoen doses per jaar.
Capaciteit die snel te vergroten is. Bij voorkeur ten minste vijf miljoen doses per jaar.
<p><span class="Smaller90"><strong>Figuur 1. </strong>Onderzoeksfasen waarin de verschillende kandidaat vaccins voor ebola zich bevinden[19]</span></p>
<p>Abstracts Najaarsvergadering 2019 NVMM en NVII<br /><span class="Smaller80">21 november 2019</span></p>
Verenigingsnieuws
2019;27:nr4
Abstracts, NVMM, Najaarsvergadering, 2019
Microbioom W.J. Wiersinga Division of Infectious Diseases, Dept. of Internal Medicine, Center for Experimental Molecular Medicine, Academic Medical Center, University of Amsterdam
Het microbioom kan beschouwd worden als een extern orgaan dat vele fysiologische functies vervult in het metabolisme, de ontwikkeling van het immuunsysteem en de afweer tegen pathogenen. Het volwassen microbioom bestaan uit 1013tot 1014 micro-organismen. De darmflora speelt mogelijk een rol bij de pathogenese van een scala aan ziektebeelden, zoals inflammatoire darmziekten, obesitas, diabetes mellitus en atopische aandoeningen. Een kanttekening hierbij is dat tot dusver voornamelijk associatiestudies uitgevoerd zijn, zonder bewijs van causaliteit. Een eenvoudige antibioticumkuur kan snel leiden tot grote verstoringen van de darmflora. Herstel van het microbioom kan maanden duren, en de nieuwe samenstelling van bacteriën kan aanzienlijk verschillen van de uitgangssituatie. Dit kan een gebruikt worden als extra argument voor het belang van antibiotic stewardship-programma’s. Dit toenemende inzicht heeft geleid tot de identificatie van nieuwe therapeutische strategieën, die op dit moment in klinische studies getest worden. Hoewel nog moet blijken wat deze kennis voor de individuele patiënt betekent, zijn verschillende interventies denkbaar, zoals suppletie van voedingsbestanddelen, prebiotica, probiotica en fecestransplantatie. Tijdens deze voordracht zal een introductie in het veld van het microbioom gegeven worden.
Effect van microbioommodulerende therapieën op verschillende ziektebeelden L. Terveer Afdeling Medische Microbiologie, LUMC, Leiden
Verscheidene microbioommodulerende therapieën passeren de revue. Van de prebiotica, probiotica en de synbiotica, naar goed gedefinieerde en gekarakteriseerde bacteriële mixen en de zeer succesvolle Feces Microbiota Transplantatie. Aan de hand van microbioom data van verschillende ziektebeelden (recidiverende Clostridium difficile, colitis ulcerosa, niet-alcoholische leververvetting en de ziekte van Parkinson) wordt meer duidelijk over het daadwerkelijke werkingsmechanisme van deze microbioommodulerende therapieën.
Het resistoom W. van Schaik Institute of Microbiology and Infection, University of Birmingham, Birmingham, Verenigd Koninkrijk
Het menselijk lichaam is gekoloniseerd door vele biljoenen micro-organismen, die collectief het humane microbioom worden genoemd. In de menselijke darm kunnen honderden verschillende bacteriële soorten worden gevonden waarvan de meeste positief bijdragen aan de menselijke gezondheid, onder andere door de productie van vitamines en de afbraak van complexe moleculen in ons voedsel. De darmen bevatten echter ook opportunistische pathogenen, zoals Enterococcus faecium en Escherichia coli. Deze bacteriën kunnen infecties veroorzaken in patiënten met een verzwakt immuunsysteem en zijn in toenemende mate resistent tegen antibiotica. Opportunistische pathogenen zijn echter niet de enige bacteriën in de darm die resistentie genen bij zich dragen want ook commensalen bevatten resistentie genen. Het totaal van antibioticumresistentiegenen in de darm wordt ook wel het resistoom genoemd. Met behulp van moderne DNA-sequentietechnologieën bestuderen wij de samenstelling van het darmmicrobioom en het resistoom tijdens ziekenhuisopname en intensieve antibiotische therapie, waarbij we opvallende verschillen en overeenkomsten vaststelden tussen patiënten in een Nederlandse en een Britse intensivecareafdeling. Ook hebben we bestudeerd in hoeverre rioolwater van ziekenhuizen en huishoudens bijdraagt aan de verspreiding van antibioticumresistentie in het milieu. Tot slot presenteer ik recente data waarin we nieuwe technologieën gebruiken om de verspreiding van antibioticumresistentiegenen in het darmmicroboom te bestuderen.
Longitudinal analysis of the gut microbiome reveals dynamic changes in relation to medications and phenotypes Jingyuan Fu1,2,*, Lianmin Chen1,2, Sanzhima Garmaeva1, Alexander Kurilshikov1, Ranko Gacesa3, Arnau Vich Vila1,3, Rinse Weersma3, Cisca Wijmenga1,*, Alexandra Zhernakova1,* 1University of Groningen, University Medical Center Groningen, Department of Genetics, Groningen, the Netherlands 2University of Groningen, University Medical Center Groningen, Department of Pediatrics, Groningen, the Netherlands 3University of Groningen, University Medical Center Groningen, Gastroenterology and Hepatology, Groningen, the Netherlands. *These authors co-directed the study
Interaction between the human genome and gut microbiome is vital to human health. While the human genome is set at birth, the gut microbiome can undergo dynamic changes over the course of an individual’s life. However, we still know little about temporal shifts in the human gut microbiome, nor about the causes and consequences of temporal shifts. We performed a longitudinal analysis on the gut microbiome of 341 participants in the Dutch population-based cohort Lifelines-DEEP, each individual having metagenomics and deep phenotypic data at two time points (~ 5 years apart). Significant temporal changes in the gut microbiome were detected at the levels of microbial composition, function and antibiotic resistance. Abundance difference was observed for 40% of taxonomies, 60% of functional pathways and 40% of antibiotic resistance genes. Furthermore, genetic stability analysis at strain level has revealed several species are under high evolution rate, including gastrointestinal disease associated Ruminococcus torques. Notable, temporal changes were associated to individual phenotypic variation and lifestyle factors. For instance, the genetic variation in Coprococcus sp ART55/1 strains was higher in individuals with larger changes in BMI, while Streptococcus thermophilus and Bifidobacterium longum showed higher mutation rates in proton pump inhibitor and non-steroidal anti-inflammatory users, as compared to non-users. Our data show that the gut microbiome sees dynamic changes not only in microbial composition and functional profiles but also in microbial genetic variation and antibiotic resistance. Our findings yield novel insights into the gut microbiome’s impact on the development of complex diseases and traits over time.
Individuele variatie in de effecten van antibiotica op darmmicrobiotasamenstelling F.H.J. Schuren, T.J. van den Broek, R.C. Montijn Microbiologie &Systeembiologie TNO Zeist
De rol die het humane microbioom speelt bij gezondheid en ziekte wordt steeds beter wetenschappelijk onderbouwd. Zowel de samenstelling als de (functionele) activiteit van het microbioom blijken een belangrijke rol te spelen, maar ook heel veel individuele variatie te vertonen. De effecten van antibioticumblootstelling op microbioomsamenstelling krijgt hierbij ook steeds meer aandacht. Binnen TNO is een platform ontwikkeld, i-screen, waarmee we darmmicrobiomen op een medium throughput wijze kunnen kweken en blootstellen aan fysiologisch relevante experimentele condities. We hebben in dit platform vijf individuele microbiotamonsters van gezonde volwassenen en 3 monsters van ouderen blootgesteld aan hoge en lage doses van 4 verschillende antibiotica: amoxicilline, clindamycine, ciprofloxacine en doxycycline. De effecten van deze eenmalige blootstellingen op de microbiotasamenstelling zijn daarna met DNA-gebaseerde methoden in kaart gebracht en nader geanalyseerd. We zien daarbij voor elk getest antibioticum andere effecten op microbiotasamenstelling, waarbij in sommige gevallen het verschil tussen volwassenen en ouderen een belangrijke rol speelt, maar in een andere gevallen de dosering de belangrijkste rol speelt. Verder zijn er grote verschillen tussen individuele monsters in de effecten van specifieke blootstellingen, waarbij sommige personen een sterke verschuiving laten zien in microbioomsamenstelling en -diversiteit, terwijl andere personen op eenzelfde blootstelling niet tot nauwelijks reageren. De consequenties van deze resultaten en hoe deze op termijn mogelijk gebruikt kunnen worden voor meer gerichte behandeling van patiënten zal nader worden toegelicht.
Pharmacokinetic-pharmacodynamic target attainment of ciprofloxacin in adult patients on general wards with adequate and impaired renal function Suzanne L. de Vroom1, Reinier M. van Hest2, Frederike V. van Daalen1, Sacha D. Kuil3, Ron A.A. Mathôt2, Suzanne E. Geerlings1, Nynke G.L. Jager2 1Department of Internal Medicine, Division of Infectious Diseases, Amsterdam Infection and Immunity (AI&II) 2Department of Hospital Pharmacy, Division of Clinical Pharmacology 3Department of Medical Microbiology, All Amsterdam UMC, University of Amsterdam, Amsterdam, the Netherlands
There are no prospective data on pharmacokinetic-pharmacodynamic (PK-PD) target attainment after the guideline-recommended dose reduction of the antibiotic ciprofloxacin in patients with impaired renal function (eGFR < 30 ml/min/1,73 m2). This study aims to investigate PK-PD target attainment of the ratio of the area under the concentration-time curve over the minimum inhibitory concentration (AUC/MIC) ≥ 125 in patients with adequate and impaired renal function receiving regular and reduced doses of ciprofloxacin. In this prospective observational cohort study adult patients on general wards of a Dutch university hospital were included when treated with ciprofloxacin. Three blood samples per patient were prospectively obtained for ciprofloxacin concentration measurement in the first 48 hours of treatment, complemented with samples from waste material. AUC calculation was performed using a population PK model developed by non-linear mixed effects modelling. A total of 40 patients were included, of which 8 patients with impaired renal function, all treated with a guideline-recommended reduced dose of ciprofloxacin. Using the clinical breakpoint MIC of most isolated bacteria (0.25 mg/L), the AUC0-24/MIC ≥ 125 was attained in 38% of patients with adequate renal function receiving a regular dose and in 13% of patients with impaired renal function receiving a reduced dose. Median drug exposure in the first 24-hours of treatment (AUC0-24) for patients with impaired renal function receiving a reduced dose was 17.9 mg/L*h, which was statistically significantly lower than the median AUC0-24 for patients with adequate renal function receiving a regular dose (29.4 mg/L*h, p < 0.01). AUC0-24/MIC ≥ 125 is not attained in the majority of adult patients on general wards for clinically relevant bacteria with MIC values at or just below the clinical breakpoint.
Data zijn eerder vermeld op 29th ECCMID in Amsterdam en op de FIGON Dutch Medicines Days 2019 in Leiden.
Surgery versus medicine in inappropriate use of intravenous and urinary catheters: a prospective observational study B.J. Laan1, M.C. Vos2, J.M. Maaskant3, M.I. van Berge Henegouwen4, S.E. Geerlings1 1Internal Medicine, Infectious Diseases, Amsterdam UMC, University of Amsterdam, Amsterdam 2Medical Microbiology and Infectious Diseases, Erasmus University Medical Centre, Rotterdam 3Clinical Epidemiology, Biostatistics and Bioinformatics, Amsterdam UMC, University of Amsterdam 4Surgery, Amsterdam UMC, University of Amsterdam
Inappropriate use of catheters is common and associated with adverse patient outcomes, such as healthcare-associated infections. We previously conducted a successful project, entitled the RICAT-study, to reduce inappropriate use of intravenous and urinary catheters in medical wards. The objective was to evaluate catheter use in surgical wards to compare this with medical wards. If there was room for improvement, we would plan to start our interventions. Inappropriate use of peripheral intravenous catheters (PIVCs) and urinary catheters was assessed in surgical wards of two university hospitals. We observed patients every other week for seven months. Inappropriate use of catheters was compared with non-surgical wards of the RICAT-study. We screened 633 patients for inclusion, of whom 409 (64.6%) were enrolled. Inappropriate use occurred in 36 (8.5%) of 425 PIVCs in 373 surgical patients, compared to 400 (22.9%) of 1747 PIVCs in 1665 medical patients. This represents a difference of 14.4% (95% CI 11.1-17.8; p < 0.001). Inappropriate use of urinary catheters occurred in 14 (10.4%) of 134 surgical patients, compared to 105 (32.4%) of 324 medical patients, a difference of 22.0% (95% CI 14.7-29.2; p < 0.001). Adjusted multivariate analyses confirmed that medical wards were independently associated with inappropriate catheter use; odds ratio of 3.50 (95% CI 2.15-5.69) for PIVCs and 2.75 (1.36-5.55) for urinary catheters. Inappropriate use of catheters is more common in medical wards compared to surgical wards. Prevention strategies should only focus on sites with high prevalence of inappropriate use.
Associatie van het gebruik van maagzuurremmers met darmkolonisatie met multiresistente micro-organismen Roel P.J. Willems1, Karin van Dijk1, Johannes C.F. Ket2, Christina M.J.E. Vandenbroucke-Grauls1 1Afdeling Medische Microbiologie en Infectiepreventie, Amsterdam UMC 2Medische Bibliotheek, Vrije Universiteit Amsterdam
Achtergrond: Maagzuurremmers worden wereldwijd veelvuldig voorgeschreven. Van maagzuurremmers is bekend dat ze de zuursecretie verminderen en de samenstelling van het microbioom van de darm verstoren. Doel: Onderzoeken van de relatie tussen het gebruik van maagzuurremmers en het risico op darmkolonisatie met BRMO’s. Opzet: Systematisch literatuuronderzoek en meta-analyse van observationeel onderzoek (case-control, cohort en cross-sectioneel onderzoek) en gerandomiseerde, gecontroleerde onderzoeken (RCTs). Methode: Wij doorzochten PubMed, Embase, Web of Science en het Cochrane Register naar artikelen gepubliceerd vóór 8 juli 2019, waarin het risico op BRMO-kolonisatie bij gebruik van maagzuurremmers werd onderzocht. Artikelen werden steeds door twee auteurs onafhankelijk van elkaar geselecteerd voor inclusie; onderzoeksgegevens werden door dezelfde auteurs onafhankelijk geëxtraheerd. Het risico op bias werd beoordeeld met behulp van de Newcastle-Ottawa schaal. Gepoolde oddsratio's werden geschat met ‘random-effect’ meta-analyse. Heterogeniteit werd onderzocht met subgroepanalyses. De PRISMA en MOOSE richtlijnen werden gevolgd en het protocol werd vooraf geregistreerd in de PROSPERO-database (CRD42018092541). Resultaten: In totaal werden 26 observationele studies (29.382 patiënten, 11.439 [39%] patiënten gebruikten maagzuurremmers) geïncludeerd; er werden geen RCT´s gevonden. Het gebruik van maagzuurremmers was geassocieerd met een bijna tweevoudig risico op darmkolonisatie met BRMO’s (ESBL, carbapenemase en/of AmpC producerende Enterobacterales en vancomycineresistente enterokokken) (OR: 1.96; 95% CI, 1.60-2.39). Resultaten werden bevestigd door sensitiviteitsanalyse van de onderzoeken die hadden gecorrigeerd voor confounding (OR, 1.74; 95% CI, 1.40-2.16). De mate van heterogeniteit was significant (72%). Conclusie: Het gebruik van maagzuuremmers leidt tot een verhoogd risico op darmkolonisatie met BRMO's. Stewardship gericht op maagzuurremmers kan mogelijk bijdragen aan preventie van BMRO-kolonisatie.
De-implementation strategy to reduce inappropriate use of intravenous and urinary catheters (RICAT): a multicentre prospective interrupted time series study B.J. Laan1, I.J.B. Spijkerman2, J.M. Maaskant3, M.J. Borgert4, M.H. Godfried4, B.C. Pasmooij4, B.C. Opmeer5, M.C. Vos6, S.E. Geerlings1 1Internal Medicine, Infectious Diseases, Amsterdam UMC, University of Amsterdam, Amsterdam 2Medical Microbiology, Amsterdam UMC, University of Amsterdam 3Clinical Epidemiology, Biostatistics and Bioinformatics, Amsterdam UMC, University of Amsterdam 4Internal Medicine, Amsterdam UMC, University of Amsterdam 5Clinical Research Unit, Amsterdam UMC, University of Amsterdam 6Medical Microbiology and Infectious Diseases, Erasmus University Medical Centre, Rotterdam
Catheter-associated bloodstream- and urinary tract infections are frequently encountered healthcare-associated infections. Our project aimed to reduce inappropriate use of intravenous and urinary catheters to prevent catheter-associated infections. In this multicentre, interrupted time series study, a de-implementation strategy with multifaceted interventions was carried out in medical wards in seven hospitals in the Netherlands. Data were collected every other week during baseline (seven months) and intervention periods (seven months). The primary outcome was the percentage of inappropriate use of peripheral intravenous catheters (PIVCs) and urinary catheters on the days of data collection. Indications for catheter use were based on international guidelines. Between September 1, 2016 and April 1, 2018, we screened 6157 patients for inclusion, of whom 5696 (92.5%) were enrolled. Inappropriate use of PIVCs occurred in 366 (22.0%) of 1665 patients in the baseline period, and 275 (14.4%) of 1912 patients in the intervention period. Primary analyses showed a relative reduction of 35% (p < 0.0001). Time series analyses showed an absolute reduction of 6.7% (95% CI 2.47 to 10.82, p = 0.011), with no significant time trends of inappropriate use of PIVCs in both periods. Inappropriate use of urinary catheters occurred in 104 (32.4%) of 324 patients in the baseline period, and 96 (24.1%) of 398 patients in the intervention period. Primary analyses showed a relative reduction of 26% (p = 0.013). Time series analyses showed no significant reduction or time trends of inappropriate use of urinary catheters. Our de-implementation strategy reduces inappropriate use of peripheral intravenous catheters in medical non-ICU patients.
Surgery versus medicine in inappropriate use of intravenous and urinary catheters: a prospective observational study B.J. Laan1, M.C. Vos2, J.M. Maaskant3, M.I. van Berge Henegouwen4, S.E. Geerlings1 1Internal Medicine, Infectious Diseases, Amsterdam UMC, University of Amsterdam, Amsterdam 2Medical Microbiology and Infectious Diseases, Erasmus University Medical Centre, Rotterdam 3Clinical Epidemiology, Biostatistics and Bioinformatics, Amsterdam UMC, University of Amsterdam 4Surgery, Amsterdam UMC, University of Amsterdam
Inappropriate use of catheters is common and associated with adverse patient outcomes, such as healthcare-associated infections. We previously conducted a successful project, entitled the RICAT-study, to reduce inappropriate use of intravenous and urinary catheters in medical wards. The objective was to evaluate catheter use in surgical wards to compare this with medical wards. If there was room for improvement, we would plan to start our interventions. Inappropriate use of peripheral intravenous catheters (PIVCs) and urinary catheters was assessed in surgical wards of two university hospitals. We observed patients every other week for seven months. Inappropriate use of catheters was compared with non-surgical wards of the RICAT-study. We screened 633 patients for inclusion, of whom 409 (64.6%) were enrolled. Inappropriate use occurred in 36 (8.5%) of 425 PIVCs in 373 surgical patients, compared to 400 (22.9%) of 1747 PIVCs in 1665 medical patients. This represents a difference of 14.4% (95% CI 11.1-17.8; p < 0.001). Inappropriate use of urinary catheters occurred in 14 (10.4%) of 134 surgical patients, compared to 105 (32.4%) of 324 medical patients, a difference of 22.0% (95% CI 14.7-29.2; p < 0.001). Adjusted multivariate analyses confirmed that medical wards were independently associated with inappropriate catheter use; odds ratio of 3.50 (95% CI 2.15-5.69) for PIVCs and 2.75 (1.36-5.55) for urinary catheters. Inappropriate use of catheters is more common in medical wards compared to surgical wards. Prevention strategies should only focus on sites with high prevalence of inappropriate use.
Contact tracing for vancomycin-resistant Enterococcus faecium (VRE): data supporting the Dutch policy of quintuple screening Linda J. Wammes1, Juliëtte A. Severin, Lonneke G.M. Bode, Margreet C. Vos, Nelianne J. Verkaik Dept of Medical Microbiology & Infectious Diseases, Erasmus MC, Rotterdam, the Netherlands 1Current affiliation: Dept of Medical Microbiology, Leiden University Medical Center, Leiden, the Netherlands
Detection of vancomycin-resistant Enterococcus faecium (VRE) transmission is hampered by low sensitivity of rectal swab cultures. Therefore, the Dutch guideline recommends screening three to five rectal swabs. We studied the number of cultures needed to detect VRE carriage in contact patients during hospital outbreaks. In a tertiary care setting, we analyzed data from nine outbreaks from 2010 until 2018. We identified 61 VRE-positive contact patients with one to five consecutive culture samples. The time between first positive culture of the presumed index patient and that of their contacts was determined, as a proxy for time needed for colonization. For 49 of 61 contact patients (80%), three or more consecutive rectal swabs were obtained. The majority of contacts (n = 39, 64%) was positive in the first culture. If only the first three rectal swabs were taken into account, 54 (89%) of VRE-positive contacts were identified. However, seven patients (11%) were found VRE-positive in the fourth or fifth culture. Moreover, time between positive culture of index and its contact was slightly shorter in contacts who were first positive in culture one to three versus those first positive in fourth or fifth culture (median (IQR) 9.0 (7-14) vs 10.5 (9.3-14.8), respectively). To detect ≥ 95% of positive contacts of VRE-positive patients, five rectal swabs were needed. The observed time between positive culture of index and contacts suggests that colonization may take several days. These findings are useful for determining the optimal VRE contact screening approach to prevent transmission in a hospital setting.
Epidemioloog, data scientist, data-analist, ...Wat is de rol van een data-expert in het medisch microbiologisch laboratorium? M.S. Berends1, S.M. Euser2, E.E.A. Hassing1, E.H.L.C.M. Hazenberg3, M. Juurlink4, Y.C. Roelofs5, M. van Rooijen6, D. Souverein2, P. Terporten7, M. Tinga8, A.F. Voor in 't holt9, I. van Weerdenburg10, J.M. Fonville11 1Stichting Certe Medische diagnostiek & advies, Groningen 2Streeklaboratorium voor de Volksgezondheid Kennemerland, Haarlem 3Jeroen Bosch Ziekenhuis, ’s-Hertogenbosch 4Isala, Zwolle 5Stichting Izore Centrum Infectieziekten Friesland, Leeuwarden 6IZ/Soa-polikliniek en streeklaboratorium GGD Amsterdam 7Medische Microbiologie, Maastricht UMC+, Maastricht 8Kenniskern Infectiepreventie, Amphia Ziekenhuis, Breda 9Medische Microbiologie en Infectieziekten, Erasmus MC, Rotterdam 10Medische Microbiologie, Radboud UMC, Nijmegen 11Stichting PAMM, Veldhoven
In medisch-microbiologische laboratoria in Nederland is in toenemende mate een vraag naar inhoudelijke data-analyse. Deze wordt uitgevoerd door de ‘data-expert’, een overkoepelende term voor functies als epidemioloog, data scientist en data-analist. We beschrijven de impact van data-gedreven medische microbiologie met recente voorbeelden van data-experts in verschillende medische microbiologie laboratoria in Nederland. Zo werd door het automatiseren van een deel van de testuitslagen de capaciteit van een SOA-polikliniek beter benut, en nam de doorlooptijd af van 66 naar 45 uur. Microbiële epidemiologische analyses hebben geleid tot aangepaste lokale empirische therapiekeuze bij sepsis. In het kader van antibiotic stewardship werd berekend dat in 11% van de gevallen is gestart met reservemiddelen zonder overleg met de arts-microbioloog, in 54% van de gevallen onterecht. Het in kaart brengen en terugkoppelen van de compliance aan bloedkweekprotocollen leidde tot een daling van 44% naar 8% enkelvoudige bloedkweken. Inzet van data-analyse en gevalideerde selectie reduceerde de tijd nodig voor registratie van postoperatieve wondinfecties, waardoor slechts 6% van de operaties nog door een medewerker beoordeeld hoeft te worden. De data-expert maakt het mogelijk om nieuwe laboratoriumprocessen te evalueren, antibioticumresistentie te monitoren, therapiekeuze te ondersteunen, spiegelinformatie terug te koppelen, kwaliteit te waarborgen, en faciliteert surveillance en infectiepreventie. Randvoorwaardelijk hieraan is het beschikken over kwalitatief goede data en een organisatie met een juist ingerichte structuur. Indien hieraan is voldaan, kan de data-expert als integraal onderdeel van de medische microbiologie een grote bijdrage leveren aan verbeterde kwaliteit van zorg, lagere kosten en klantenbinding.
Inhoud van de cursus Van 15 tot en met 17 mei 2019 vond in het Amsterdamse Hotel Casa de tweejaarlijkse serologiecursus plaats. De cursus wordt georganiseerd door de stichting Serology Education, die bestaat uit artsen-microbioloog en arts-assistenten uit het VUmc en het Elisabeth-TweeSteden Ziekenhuis. Het is een groot opgezette cursus met circa 150 deelnemers, grotendeels aios medische microbiologie is, maar ook artsen-microbioloog, analisten en nog enkele andere geïnteresseerden. Het thema van de eerste dag is ‘laboratoriumzaken’, waaronder basistechnieken in de serologie, basisimmunologie en kwaliteit. Op de tweede dag staan zwangeren, immuungecompromitteerden en reizigers centraal. Deze dag wordt onder andere ingegaan op CMV, TORCHES, gisten en schimmels. Op de derde dag komen specifieke infecties aan bod, en passeren onder andere hepatitis A t/m E, vaccinaties bij immuungecompromitteerden, Mycobacterium tuberculosis, Borrelia en lues de revue. De vrijdag wordt afgesloten met een zogeheten Kahoot-quiz, waarin alle deelnemers het online tegen elkaar kunnen opnemen. Via de Kahoot-app kunnen ze per vraag op het juiste antwoord 'stemmen'.
Review Deze serologiecursus is weliswaar niet verplicht maar wordt wel als een onmisbare cursus gezien door veel arts-assistenten. Aangezien deze cursus maar eenmaal in de twee jaar wordt gegeven, zijn er veel deelnemers. Desondanks voelt het niet massaal. De cursuslocatie is prettig en ruim. In de pauzes is er voldoende gelegenheid om andere deelnemers en de organisatie te spreken, evenals tijdens het cursusdiner op donderdagavond.
Tijdens de voordrachten is er voldoende ruimte voor interactie, waar ook regelmatig gebruik van wordt gemaakt. Dit wordt versterkt door het veelvuldig gebruik van de Kahoot-app. Als de vrijdagmiddag wordt ingeluid door de Bring Sally Up Squat challenge is het ijs definitief gebroken en is iedereen klaar voor een laatste middag aan onderwijs. Inhoudelijk wordt een breed palet aan infectieziekten en serologische bepalingen besproken. Geheel begrijpelijk wordt daarmee vaak eerst ingegaan op de infectieziekte zelf en daarna op de betreffende diagnostiek. Dit geeft de voordrachten een meer klinische oriëntatie, maar dit gaat soms wel enigszins ten koste van de diepgang van de technische details.
Samengevat: de serologiecursus is een zeer leuke en interessante cursus, die een goed overzicht biedt van de serologie in de breedste zin van het woord, en waarin de bacteriologie, virologie, mycologie en parasitologie uitgebreid aan bod komen.
25 oktober 2019 - R.M. Koch Differential pathogen-host interactions
Promotor: prof. dr. P. Pickkers Copromotoren: dr. M. Kox en dr. M.I. de Jonge Radboudumc Nijmegen, afd. Intensieve Zorg
18 november 2019 - M.G. Tapia Calle Human cell-based in vitro system for vaccine evaluation
Promotoren: prof. dr. prof. dr. A.L.W. Huckriede en prof. dr. C.A.H.H. Daemen UMC Groningen, afd. Medische Microbiologie
22 november 2019 - H.R.A. Streefkerk Electronically assisted surveillance of hospital-acquired infections
Promotor: prof. dr. H.A. Verbrugh (emeritus) Copromotor: dr. R.P. Verkooijen Erasmus MC Rotterdam, afd. Medische Microbiologie & Infectieziekten
27 november 2019 - S. Nepal A proteogenomic view on antibiotic resistance in pathogenic Enterobacter species
Promotoren: prof. dr. J.M. van Dijl en prof. dr. J.W.A. Rossen Copromotor: dr. D. Bathoorn UMC Groningen, afd. Medische Microbiologie en Infectiepreventie
4 december 2019 - A. Boerekamps Hepatitis C infection in Dutch HIV-positive patients in the era of direct-acting antiviral therapy
Promotor: prof. dr. A. Verbon Copromotor: dr. B.J.A. Rijnders Erasmus MC Rotterdam, afd. Medische Microbiologie & Infectieziekten en Sectie Infectieziekten, afd. Inwendige Geneeskunde
6 december 2019 - I. Taufik Mechanism of Translocon
Promotor: prof. dr. A.J.M. Driessen Rijksuniversiteit Groningen, Moleculaire Microbiologie. Groningen Biomolecular Sciences and Biotechnology Institute
10 december 2019 - R.A. Wijma Pharmacokinetic profiling of fosfomycin and nitrofurantoin to optimize the treatment of uncomplicated urinary tract infections
Promotoren: prof. dr. J.W. Mouton† en prof. dr. T. van Gelder Copromotor: dr. B.C.P. Koch Erasmus MC Rotterdam, afd. Medische Microbiologie & Infectieziekten en afd. Apotheek
11 december 2019 - M.J.C. van den Beld Shigella spp. and entero-invasive Escherichia coli
Promotor: prof. dr. J. W.A. Rossen Copromotoren: dr. A.M.D. Kooistra-Smid, dr. F.A.G. Reubsaet UMC Groningen, afd. Medische Microbiologie
11 december 2019 - E.F. Lizarazo Forero Epidemiology, genetic diversity and clinical manifestations of arboviral diseases in Venezuela
Promotoren: prof. dr. J. W.A. Rossen en prof. dr. A.W. Friedrich Copromotoren: dr. A. Tami en prof. dr. M.E. Grillet UMC Groningen, afd. Medische Microbiologie
7 januari 2020 - S. Colombe HIV and Schistosoma spp. interactions: epidemiology and consequences for detection and prevention in the lake region of Tanzania
Promotor: prof. dr. M. Yazdanbakhsh LUMC Leiden, afd. Parasitologie
7 januari 2020 - A.F. Muggen Chronic Lymphocytic Leukemia: the B cell receptor and beyond
Promotoren: prof. dr. J.J.M. van Dongen en prof. dr. R.W. Hendriks Copromotor: prof. dr. A.W. Langerak Erasmus MC Rotterdam, afd. Immunologie en afd. Longziekten
13 januari 2020 - M.C.L. Böger Structure-function relationships of (prebiotic) carbohydrates and their selective consumption by probiotic bacteria
Promotor: prof. dr. L. Dijkhuizen Rijksuniversiteit Groningen, Microbiële Fysiologie. Groningen Biomolecular Sciences and Biotechnology Institute
20 januari 2020 - A.C. da Cruz Campos Comprehensive characterization of Escherichia coli isolated from urine samples of hospitalized patients in Rio de Janeiro, Brazil
Promotor: prof. dr. J. W.A. Rossen Copromotor: dr. M.A. Chlebowicz-Flissikowska UMC Groningen, afd. Medische Microbiologie
24 januari 2020 - B.M.E. Cahueñas Effects of air pollution on the respiratory health and the respiratory immune system. Studies in Ecuadorian Children
Promotor: prof. dr. P.D. Katsikis Copromotor: dr. ir. T. Voorman Erasmus MC Rotterdam, afd. Immunologie
AFSCHEIDSREDE
17 januari 2020 - prof. dr. H.V. Westerhoff Afscheidsrede van prof. dr. H.V. Westerhoff als hoogleraar Molecular Cell Physiology, met als titel: “Trumping the Trumps: addressing complexities through biological rationality.” VU Amsterdam, Sectie Moleculaire Celfysiologie.